Experimental Testing of the Role of Cytoskeleton in the Solution by Neurons of Problems Facing the Brain
E. A. Liberman1, S. V. Minina1, D. A. Moshkov2, I. M. Santalova2, I. A. Chistopolskiy3, and N. E. Shklovski-Kordi4*
1Institute of Information Transmission Problems, Russian Academy of Sciences, B. Karetnyi Pereulok 19, 101447 Moscow, Russia; fax: (495) 209-0579; E-mail: efim@liberman.ru; svminina@mail.ru2Institute of Experimental and Theoretical Biophysics, Russian Academy of Sciences, 142290 Pushchino, Moscow Region, Russia; fax: (496) 733-0553; E-mail: santalova@iteb.ru
3Koltsov Institute of Developmental Biology, Russian Academy of Sciences, ul. Vavilova 26, 119991 Moscow, Russia; fax: (495) 135-8012; E-mail: compar@pochtamt.ru
4Hematology Research Center, Russian Academy of Medical Sciences, Novozykovskii pr. 4a, 125167 Moscow, Russia; fax: (495) 438-1800; E-mail: nikitashk@gmail.com
* To whom correspondence should be addressed.
Received June 27, 2007; Revision received September 14, 2007
Investigation of the influence of cAMP on neuronal electric activity suggests that nerve cells can solve problems using an intraneuronal calculating medium based on the cytoskeleton. When a new problem is posed, this structure has to be disassembled and assembled by the neuronal molecular computer according to the program recorded in DNA. If DNA lacks an appropriate program, the cytoskeleton will not be assembled. In our experiments, fishes which were rotated simultaneously around two mutually perpendicular axes lost their swimming ability, and some dramatic changes were observed in the cytoskeleton of their Mauthner neurons. These changes disappeared after a long-term rest: the cytoskeleton was restored simultaneously with the ability for normal swimming.
KEY WORDS: molecular quantum generator, cytoskeleton, calculating mediumDOI: 10.1134/S0006297908040147
Abbreviations: MC) molecular computer; MN) Mauthner neurons of fishes.
For a long time biochemical processes inside a neuron were considered to
be a source of energy for maintaining an ion gradient on the outer
membrane required for the principal function of the neuron, i.e.
generation of nerve impulses. According to this concept, generation of
impulses is regulated only by external molecules (mediators), which
open or close ion channels of the neuron outer membrane on binding with
external receptors.
But a hypothesis about a molecular computer (MC) of the cell [1] stimulated studies on intracellular processes involved in information processing by the neuron. According to this hypothesis, the living cell is controlled by a parallel-serial stochastic molecular machine, which can be equivalent to a universal computer. The MC operates with definitely addressed molecule-words. Molecule-words are processed by molecular operators with appropriate addresses. Molecule-words enter contacts with the corresponding molecular operators as a result of interaction between complementary moieties of molecules during Brownian motion.
The RNA processing discovered later [2] completely fit this scheme.
However, a computer operating with a DNA-RNA-protein system and using Brownian motion to search for addresses acts slowly. It was obvious that it was unable to solve physical problems of the whole body behavior, and a hypothesis was proposed of a neuronal quantum molecular regulator [3] using the cytoskeleton as a calculating medium [4].
This hypothesis was partly based on data on the influence of cAMP (a substance synthesized inside the cell) on electric activity of the neuron [5]. It has been shown that the intracellular injection of cAMP as it is, without a mediator, induces depolarization of the nerve cell membrane [6, 7]. This depolarization is conditioned by opening of sodium and closing of potassium channels of the neuronal membrane [8]. The effect of cAMP was markedly decreased upon destruction of the neuronal cytoskeleton caused by incubation of the ganglion at low temperature in the presence of nocodazole [9]. A mechanical irritation of neurons by injections under pressure of large portions (resulting in a noticeable increase in the cell volume) of indifferent solutions also induced a generated potential [10].
According to the hypothesis about the intracellular processing of information to provide for the ability of the neuron to solve different problems, the cytoskeleton has to be disassembled and reassembled by the neuronal molecular computer according to the program recorded in DNA. If there is no appropriate program, the cytoskeleton will not be assembled. Such conditions could be expected if an unsolvable problem faced an animal. Such a problem was exemplified by a simultaneous rotation of the animal's body around two mutually perpendicular axes. The first achievements in this field are reported in the present paper.
MATERIALS AND METHODS
Mauthner neurons (MN) of fishes are the most suitable object for studies on the role of the cytoskeleton. They are the only identified twin neurons of vertebrates which control tail movement in fishes and amphibians. In experiments 110 goldfishes (Carassius auratus shubunkin) of the same hatch at the age of three-four months and of about 3 cm in length were used. The control group of fishes was not subjected to rotation. The experimental fishes were rotated simultaneously in two mutually perpendicular directions for 2 h. For electron microscopy, MN-containing fragments of the myelencephalon were isolated, fixed, and treated as described in [11]. Ultrathin sections containing the soma and dendrites of MN were analyzed using a TESLA BS-500 electron microscope at magnification of 10,000.
RESULTS
Before the experiment, the fishes actively swam in the whole space of the aquarium. After simultaneous rotation for 2 h around two mutually perpendicular axes at the rate of 30 rpm, the fishes lost the ability to swim, and within an hour or more they lay on their bellies motionless on the aquarium bottom. Upon resting for 24 h, the fishes reacquired their ability to swim.
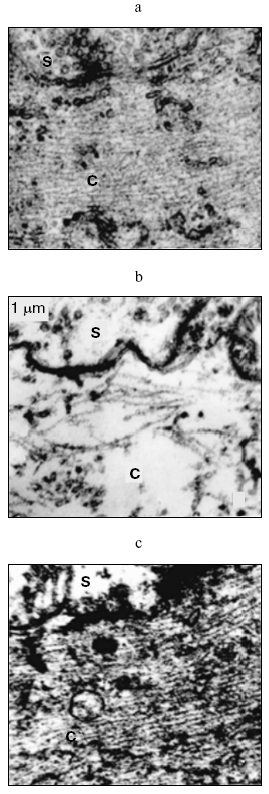
Ultrastructural changes in MN are shown in the figure. Intact MN are characterized by an ordered cytoskeleton of closely disposed microfilaments parallel to the major axis of the dendritis with a regular distance between them (figure, panel (a)). After the experimental exposure, the MN of the fishes manifested substantial damage to the cytoskeleton presented by a sharply pronounced loosening of its network. The microfilaments lost their ordered orientation, became short and twisted, and the distance between them increased (figure, panel (b)). Upon the rest for 24 h, the fishes restored their capability for swimming, and the MN cytoskeleton renormalized (figure, panel (c)).
Introduction into the water of gel capsules allowed us to reduce mechanical damage to the animals during the rotation, but the specific destruction of the cytoskeleton shown in the figure remained.Ultrastructure of goldfish MN: a) intact MN; b) MN after rotation for 2 h around two mutually perpendicular axes; c) MN after 24 h of rest. C, cytoplasm; S, synapse
DISCUSSION
The quantum molecular computer of neurons was earlier supposed to consist of the intracellular cytoskeleton used as a molecular calculating medium. The channels, which are opened upon activation by a mediator, send inward a high frequency quantum-mechanical signal. Hypersonic phonons generated by the ion flux through the entrance channels are distributed over the cytoskeleton network. Interference, absorption, and generation of new phonons at the cost of heat movement and free energy of biochemical reactions contribute to solution of problems facing the cell. The program for solution of these problems is recorded in DNA and is realized at the cost of cytoskeleton assembly.
According to this idea, the neuron has entrance channels controlled by receptors and exit channels controlled by the cAMP system [4]. The molecular calculating medium connects the entrance ion channels decoding the synaptic information to sonic signals with the exit channels, which are controlled by sonic signals transformed by the cytoskeletal network. Sonic waves distributing across different calculating media are responsible for solution of different physical problems.
When describing a living organism, one has to take into account that the cells are molecular quantum computers for which the influence of measurement and calculation on the problem under solution is essential. To describe such systems, it is necessary to create a new science based on four principles as follows: principle of lowest price for measurement and calculation; principle of optimality; principle establishing that natural law irreversibility caused by the influence of measurements and calculations; and newly formulated principle of causality [12, 13].
Contemporary physics describes the world using the principle of relativity. This principle asserts that all coordinate systems are equivalent. The major idea of the new science discriminating it from contemporary physics is that not only influence of the measurement which has determined the arising of quantum mechanics, but also influence of the calculation on the problem under solution are essential for a living organism [1, 13]. The principle of relativity for living systems is only a special case of the optimality principle. The optimality principle states that for complicated systems there is a selected coordinate system providing for the solution of a problem without the influence of measurements and calculations on the problem itself.
In fact, the equivalence of laws in all coordinate systems is very convenient for living organisms. This allows us to walk using coordinates of stationary walls to solve problems in our brain and personal self-consciousness. But if a human begins to rather rapidly rotate on a place, the walls begin to move in his consciousness. This effect is usually spoken of as “a human has their head swimming”. To scientifically describe this generally known phenomenon, we have supposed that the human's neurons and personal self-consciousness pass into the coordinate system rotating together with him [4]. It seems that the difficulty of calculation to solve the problem of maintaining the body balance inside the system of stationary walls becomes so complicated that it cannot be solved without the influence of calculation on the problem under solution. Therefore, the problem of a ballerina executing thirty-two fouetté or of an athlete performing somersaults is to twist the environment around themselves as soon as possible. We think that DNA already has programs determining the cytoskeletal structure to provide for adequate human behavior in the coordinate system rotating together with their body. These programs are to be more intricate than programs of general relativity theory. According to our hypothesis, a comparatively rapid transition into a rotating coordinate system occurs at the cost of a corresponding change in the cytoskeleton inside the neutrons controlling these movements. This change can be realized at the cost of a process similar to muscle contraction, possibly by means of different types of myosins in nerve cells, i.e. in response to calcium entrance through special channels.
The first experimental testing of the optimality principle was performed on a human in an aerospace laboratory [14]. The subject was rotated around a vertical axis of his body and then asked to rapidly show with his hand where a target appeared for a short time. Despite the action of the Coriolis force on the subject's hand, all the subjects made virtually no mistakes, because their brain gave onto their muscles a signal “to make a corresponding correction”. But when a virtual reality helmet was put on the subjects and their consciousness believed that they were rotating, all of them indicated the target with mistakes because their brain sent to the hand muscles a correction for the lacking Coriolis force.
In addition to receptors sending signals for a fast rearrangement of the cytoskeleton to allow the motor neuron to rapidly pass to a state required for solution of the problem within the coordinate system rotating together with the animal's body, receptors should exist responsible for sending signals for the necessity of a substantial rearrangement of the cytoskeleton to solve novel problems. The cytoskeleton has to be disassembled and reassembled by the neuronal molecular computer according to the program recorded in DNA. If DNA has no appropriate program, the cytoskeleton will not be reassembled. Such conditions are expected to arise if an insoluble problem faces an animal. Such a problem was exemplified by a simultaneous rotation of the animal's body around two mutually perpendicular axes. Note, that from the viewpoint of physics this problem can be easily solved. Only the influence of measurement and calculation makes it insoluble.
Obviously, in experiments on humans it is impossible to find out what is really happening in the brain neurons on transition into the coordinate system rotating together with the body. However, it has been demonstrated in the above-described experiments on animals. Working along this line, we hope to obtain direct experimental confirmation of the optimality principle.
The new science cannot answer the question of how the world inhabited by us is constructed, but it successfully responds to another question which is more important for humans: why the world is such as it is, for which purpose it is quantized and wave. Quantum and wave properties provide for the existence of stable atoms, molecules, solid states, fluids, and gases. For complete comprehension, we have also to answer another question: what for a single phonon examines the intracellular structure? And there is the following answer: to provide for an efficient operation of quantum molecular regulators of living cells, which use the cytoskeleton as a calculating medium.
This work was supported by the Russian Foundation for Basic Research (project No. 06-04-48898).
REFERENCES
1. Liberman, E. A. (1972) Biofizika, 17, 932-943.
2. Crick, F. (1979) Science, 204, 264-271.
3. Liberman, E. A. (1983) Biofizika, 28, 183-186.
4. Minina, S. V., and Liberman, E. A. (1990) Biofizika, 35, 132-134.
5. Liberman, E. A., Minina, S. V., and Shklovski-Kordi, N. E. (1989) Biosystems, 22, 135-154.
6. Liberman, E. A., Minina, S. V., and Golubtsov, K. V. (1975) Biofizika, 20, 451-456.
7. Liberman, E. A., Minina, S. V., and Golubtsov, K. V. (1977) Biofizika, 22, 75-81.
8. Avdonin, V. B., Liberman, E. A., Liberman, A. E., Minina, S. V., and Myakotina, O. L. (1999) Biofizika, 44, 75-82.
9. Liberman, E. A., Minina, S. V., Myakotina, O. L., and Shklovski-Kordi, N. E. (1985) Biofizika, 30, 632-636.
10. Liberman, E. A., Minina, S. V., Myakotina, O. L., Shklovski-Kordi, N. E., and Conrad, M. (1985) Brain Res., 338, 33-44.
11. Santalova, I. M., and Moshkov, D. A. (1999) Neuroscience, 89, 593-602.
12. Liberman, E. A. (1997) Biofizika, 42, 988-991.
13. Liberman, E. A., Minina, S. V., and Shklovski-Kordi, N. E. (2001) Biofizika, 46, 765-767.
14. Lackner, J. R., and DiZio, P. (2000) Exp. Brain Res., 130, 2-26.