Localization of Mullerian Inhibiting Substance Receptors in Various Human Cancer Cell Lines
A. V. Rodina*, N. V. Gukasova, V. A. Makarov, I. G. Kondrasheva, A. V. Khomyakova, G. A. Posypanova, O. N. Popova, E. Yu. Moskaleva, and S. E. Severin
Moscow Research Institute of Medical Ecology, Moscow Department of Health Care, Simferopolsky blvd. 8, 117638 Moscow, Russia; fax: (499) 613-4818; E-mail: sergsev@aha.ru; allrodina@yandex.ru* To whom correspondence should be addressed.
Received January 15, 2008; Revision received March 7, 2008
Recombinant human MIS (rhMIS) produced in transfected Chinese hamster ovary cells has been purified by immunoaffinity chromatography. In the absence of reducing agents, 140 kD homodimer and several oligomers with molecular masses from 280 to 1000 kD are present. Homodimer, tetramer, and higher-molecular-weight rhMIS fractions reduced survival of tumor cells. For these experiments, FITC-labeled rhMIS was used for binding and endocytosis studies by flow cytometry. Flow cytometry performed on MIS-sensitive cancer cell lines demonstrated specific binding of rhMIS. The majority of rhMIS receptors have cytosolic localization. Thus, the level of MIS receptors on the cell membrane was proportional to the content of MIS-binding proteins in the whole cell and defines a level of receptor-mediated endocytosis. The immunopurified rhMIS caused significant growth inhibition of ovarian and prostate adenocarcinoma and melanoma human cell lines in inhibition assays.
KEY WORDS: human recombinant MIS, receptor localization, biological activityDOI: 10.1134/S0006297908070080
Abbreviations: BSA) bovine serum albumin; DAB) 3,3´-diaminobenzidine; DMSO) dimethyl sulfoxide; FCS) fetal calf serum; FITC) fluorescein isothiocyanate; mAB) monoclonal antibody; MIS) Mullerian inhibiting substance; MISR II) MIS receptor type II; PBS) phosphate-buffered saline; PBS-T) PBS containing 0.5% Tween-20; PF) paraformaldehyde; PMSF) phenylmethylsulfonyl fluoride; rhMIS) recombinant human MIS.
The MIS (Mullerian inhibiting substance) protein, which was first
discovered in 1947 by A. Jost as an inducer of Mullerian duct
regression in embryogenesis, possesses a broad spectrum of activity
being involved in inhibition of oocyte and spermatogonium maturation,
in differentiation of gonads, and regulation of fetal lung development.
An important function of this protein is suppression of tumor growth,
in particular due to inhibition of epidermal growth factor receptor
autophosphorylation [1].
MIS is a member of the TGFbeta superfamily and, like all proteins of this family, such as TGFbeta, activins, and BMP, mediates biological effects via binding with its transmembrane type II receptor (MISR II), which, after binding with MIS, forms a heterodimer complex with the type I receptor (MISR I). This is followed by transphosphorylation of MISR I by the MISR II receptor at a GS-site localized in the cytoplasm close to the transmembrane region. Activation of MISR I leads to phosphorylation of receptor-specific Smad-proteins, which in turn bind with the Smad-4 coactivator; this complex is translocated into the nucleus and participates in regulation of gene expression [2, 3]. The specific MISR II receptor is conservative, whereas MISR I receptors, also known as activin receptor-like kinases (ALKs), are different depending on tissue context [4-8].
MIS causes apoptosis of cells expressing MISR II receptors [9-13]. However, the study of mechanisms of programmed death of cells providing regression of Mullerian duct has demonstrated that MISR II receptors are expressed in mesenchymal cells surrounding the duct, whereas the apoptotic cells are found in epithelium, thus implying paracrine, as well as autocrine, mechanism of MIS action [14].
It was determined that more than 50% of cells from ascitic fluid of patients with ovarian carcinoma stages III and IV expressing MISR II mRNA bound the recombinant human MIS (rhMIS), and rhMIS inhibited their proliferation in experiment ex vivo [9]. It was also demonstrated that rhMIS inhibits, both in vitro and in vivo, the proliferation of eye melanoma cells of neuronal genesis [15]. The level of MIS receptor expression by various cells may be of great prognostic importance for determination of their sensitivity to this protein when it is therapeutically applied.
In connection with this, the goal of our study was characterization of human tumor cells of various geneses, including cells of skin melanoma, by amount and localization of MIS receptors and by capability of binding and endocytosis of this protein in comparison with sensitivity of the examined cells to the action of rhMIS.
MATERIALS AND METHODS
Isolation of rhMIS from the culture medium of CHO cells. The Chinese hamster CHO line cells (kindly provided by P. K. Donahoe) transfected with human MIS gene cDNA and secreting this protein into the culture medium were grown in plastic rollers (Corning-Costar, The Netherlands) in ribo- and deoxyribonucleotide-free alphaMEM medium (Invitrogen Life Technologies, USA) containing 100 ng/ml of methotrexate (Ebewe, Austria), 5% fetal calf serum (FCS) (Hy Clone, USA), 50 µg/ml of gentamicin (ICN, USA), and 1% L-glutamine. The cultural medium of the CHO cells was collected, concentrated using an Amicon concentrator cell (membrane PM-10; Millipore, USA), and then phenylmethylsulfonyl fluoride (PMSF) (final concentration 1 mM) and NaN3 (final concentration 0.04%) were added. The medium was centrifuged for 20 min at 4°C and 3000g, and the supernatant (950 ml) was applied onto an immunoaffinity column (5 × 1.6 cm) with affinity sorbent prepared using the monoclonal antibody (mAB) raised against rhMIS (clone 6E11; provided by State Research Institute of Highly Pure Biopreparations, St. Petersburg, Russia). The affinity sorbent (5.5 ml) was equilibrated with 50 ml of 0.02 M Hepes, pH 7.4, at 4°C. Then the supernatant was applied onto the column (flow rate 1 ml/min) at 4°C. After application, the column was washed with 50 ml of 0.02 M Hepes, pH 7.4, at 4°C and then with 50 ml of 0.5 M NaCl in 0.02 M Hepes, pH 7.4, at room temperature. The protein was eluted with 0.1 M Gly·HCl, pH 2.5, and neutralized with 1 M sodium phosphate, pH 8.0, under agitation. The neutralized eluate was dialyzed against phosphate-buffered saline (PBS), pH 7.4, for 18 h at 4°C. The dialyzate was centrifuged at 3000g for 20 min at 4°C (BR 4i centrifuge; Jouan, France). The supernatant was concentrated using an Amicon concentrator cell. The concentration of MIS was determined by sandwich-ELISA. To perform ELISA, 96-well plates (Corning-Costar) were sensitized with anti-MIS mAB (clone 6E11, 5 µg/ml), the solid-phase free surface was blocked with 1% bovine serum albumin (BSA) solution (ICN), followed by addition of samples containing MIS, then the 6E11 mAB-horseradish peroxidase conjugate (5 µg/ml) was added followed by addition of the substrate, 3,3´,5,5´-tetramethylbenzidine. The reaction was terminated by addition of 0.05 M H2SO4, and the optical density at lambda = 450 nm was measured on a Multiskan MS plate spectrophotometer (Labsystem, Finland).
The purity and homogeneity of MIS protein were analyzed by electrophoresis in polyacrylamide gel according to the method of Laemmli using 4 to 15% gel gradient under reducing and non-reducing conditions (reduced samples were treated with 0.1 M beta-mercaptoethanol) and by gel filtration on a Superdex 200 HR column (Amersham Pharmacia Biotech, USA). Protein standards in the range of 20-97 kD were used for molecular mass determination.
Immunochemical characterization of rhMIS using immunoblotting. The rhMIS samples were separated in polyacrylamide gel according to Laemmli (4-15% gradient under non-reducing conditions) followed by electrotransfer on a PVDF membrane (Immobilon-PSQ Transfer Membrane; Millipore) in buffer containing 39 mM glycine, 48 mM Tris, 0.037 mM SDS, and 20% methanol using a Hoeffer SemiPhor device for semidry blotting (Amersham Pharmacia Biotech). The membrane was then incubated in blocking buffer (5% nonfat-dried milk in PBS-T), washed twice with PBS-T (PBS containing 0.5% Tween-20), and incubated with anti-MIS mAB (clone 6E11, 5 µg/ml) followed by addition of rabbit antibodies against mouse immunoglobulins conjugated with horseradish peroxidase (working dilution 1 : 1000). To visualize proteins, substrate mixture containing 0.025% 3,3´-diaminobenzidine (DAB), 0.075% 4-chloro-1-naphthol, and 0.01% H2O2 was used.
Cell growth. The OVCAR8 and MOVCAR7 cells were kindly provided by P. K. Donahoe. The human ovarian carcinoma line OVCAR8 cells (the cells expressing MISR II receptors [10]) and murine ovarian carcinoma line MOVCAR7 cells (the cells transfected with the MISR II receptor gene [12]) were grown in plastic culture flasks (Corning-Costar) in DMEM medium (ICN) containing 10% (OVCAR8) or 4% (MOVCAR7) FCS (Gibco, USA) and 50 µg/ml of gentamicin (ICN) in a CO2 incubator (Jouan) at 37°C in humid atmosphere containing 5% CO2. The medium for MOVCAR7 growth was supplemented with insulin-transferrin-sodium selenite (1 : 500; Gibco). The human ovarian carcinoma line SKOV3 cells (negative control; in this line the mechanism of signal transduction from MISR II is impaired [16]) and SKVLB (selected from the SKOV3 line by resistance to vinblastine [17]) were grown under the same conditions, but RPMI 1640 medium (ICN) containing 10% FCS was used. The prostate carcinoma line LNCaP cells were grown in medium representing the mixture of RPMI 1640 (Sigma, USA) and DMEM (Sigma) media in the ratio 1 : 1 (v/v) supplemented with 10% FCS and 5% horse serum (Gibco). The cells of prostate carcinoma line Du145 and melanoma line MS were grown in DMEM medium containing 10% FCS. The cells were passed twice a week and picked off the plastic using either a Versene solution (PanEko, Russia) (SKOV3, SKVLB, LNCaP, Du145, and MS) or 0.05% trypsin (Serva, Germany) in Versene solution (OVCAR8 and MOVCAR7).
Preparation and growth of primary lines of human melanoma cells. Melanoma fragments were taken from tumors of surgery patients, connective tissue was removed, the tumor tissue was disintegrated, and the cells were suspended using a glass homogenizer. Then the cell suspension was washed with culture medium by centrifugation, resuspended in complete culture medium containing the RPMI 1640 (Sigma) and DMEM (Sigma) media taken at the ratio 1 : 1 and supplemented with 10% FCS up to the final concentration of 0.5·106 cells/ml, and placed in culture flasks (Corning-Costar), 5 ml in each flask. After attachment of tumor cells and removal of unattached cells, the cells were grown in complete culture medium, passing twice a week, and picking up from the substrate using the Versene solution.
Determination of antiproliferative activity of MIS using the MTT-test. The cells were seeded into the wells of a 96-well plate (Corning-Costar), 2-4 thousand cells in a well (in 200 µl), one day before the experiment. MIS at varied concentration was added, and the cells were incubated under standard conditions of growth for 72-144 h. Cell viability was determined using the MTT-test according to the method of Mosmann [18]. Four hours before the incubation finish, 50 µl of MTT (Sigma) solution at concentration of 1 mg/ml of the culture medium was added into each well. After color development, the medium was removed, the sedimented formazan crystals were dissolved in 100 µl of dimethyl sulfoxide (DMSO), and the color intensity was measured by absorption at 540 nm on a plate spectrophotometer (Labsystem). The cell survivability was estimated as percentage of untreated control.
Synthesis of fluorescein isothiocyanate (FITC)-labeled MIS. Five microliters of FITC solution in DMSO (1 mg/ml) was added under agitation to 700 µg MIS in 1 ml of 0.1 M NaHCO3, pH 8.8. The reaction mixture was agitated for 2 h at room temperature and overnight at 4°C and placed onto a column (1 × 28 cm) with Sephadex G-25f equilibrated with PBS, pH 7.4. The protein was eluted with the same buffer (elution rate 1 ml/min), and the elution profile was monitored by absorption at 280 nm. The first fraction (about 5 ml) was collected and concentrated on an Amicon cell with PM-10 membrane to 300 µl. The protein concentration was measured using the BCA set (Sigma). Concentration of FITC was determined by spectrophotometry assuming the molar extinction coefficient of FITC epsilon498 = 68,000 M-1·cm-1. The MIS/FITC ratio was 1 : 2.
Examination of MIS binding and endocytosis by flow cytofluorometry. Before experiment, the cells were incubated for 1 h in medium devoid of serum. To analyze the binding, MIS-FITC in the concentration range of 100-1000 nM was added to the cell suspension in the cold and incubated at 4°C for 1 h. When accumulation of MIS-FITC was examined, the sample was incubated at 37°C. One sample contained 2·105 cells in 200 µl of PBS containing 1% BSA. After finishing the incubation, the cells were washed thrice with cold PBS containing 0.1% BSA and fixed with 2% paraformaldehyde (PF) in PBS. Fluorescence intensity was measured using an EPICS-XL flow cytofluorometer (Beckman Coulter, USA). Fluorescence was excited by an argon laser (lambdaex = 488 nm, emission bandwidth 515-520 nm).
Evaluation of MIS-FITC binding level in different tumor cell lines after intracellular staining. Intracellular staining was performed using FITC-labeled MIS. Before experiment, the cells were incubated for 1 h in medium devoid of serum. Then the cells were washed twice with washing buffer (PBS containing 1% BSA and 0.1% NaN3) and fixed with 4% PF for 30 min at room temperature. Following fixation, the cell preparations were washed twice with the washing buffer and treated for 5 min with the same buffer containing 0.5% saponin for permeabilization of cell membranes. After permeabilization, MIS-FITC at concentration of 70 µg/ml was added to the cell suspension followed by incubation for 1 h at 4°C. Following two washes with PBS containing 0.1% BSA, 0.1% saponin, and 0.1% NaN3, the cells were fixed with 1% PF for 30 min at room temperature and fluorescence intensity was measured on the EPICS-XL flow cytofluorometer. To estimate specificity of intracellular binding of MIS-FITC, the permeabilized cells were treated with 1.4 mg/ml of rhMIS (20-fold excess), fixed with 1% PF for 30 min, and washed twice with PBS containing the above-mentioned additives. Then MIS-FITC at the concentration of 70 µg/ml was added to the cell suspension. The cells were washed twice, fixed with 1% PF, and fluorescence intensity was measured.
RESULTS
Characterization of rhMIS preparation. The purity and homogeneity of the rhMIS preparation isolated from the culture medium of CHO cells by immunoaffinity chromatography were tested using PAGE. Figure 1a demonstrates a typical electrophoregram of rhMIS preparation under reducing (lane II) and non-reducing (lane III) conditions. A single band with molecular mass about 70 kD is found under reducing conditions, whereas under non-reducing conditions four bands are visible, thus indicating the presence of protein fractions with molecular mass of 140 (4), 280 (3), and more than 1000 kD (1 and 2).
It has been demonstrated using immunoblotting (Fig. 1b) that all four bands found under non-reducing conditions are stained with anti-MIS antibody. This indicates that the rhMIS preparation contains a MIS homodimer with molecular mass of 140 kD as well as oligomeric structures stabilized by disulfide bonds.Fig. 1. Characterization of rhMIS preparation. a) Polyacrylamide gel electrophoresis of rhMIS preparation under reducing (II) and non-reducing (III) conditions. I, protein standards. b) Immunoblotting of rhMIS preparation after electrophoresis under non-reducing conditions (I). II, protein standards. c) Photometric elution profile (280 nm) for rhMIS after gel filtration on a Superdex 200 HR column. Areas under peaks: 1, 2) 29.09%; 3) 29.64%; 4) 32.85%. The typical results of one of four experiments are given.
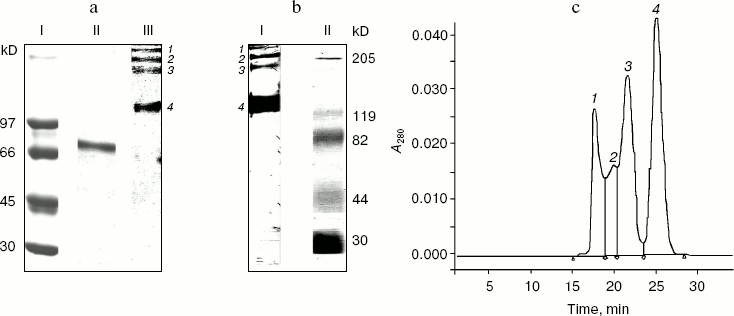
Gel filtration of the rhMIS preparation on the Superdex 200 HR column revealed four protein peaks (Fig. 1c), which corresponds to the data of electrophoresis and immunoblotting. For examination of biological activity, protein fractions 3 and 4 corresponding to analogous fractions found after electrophoresis were collected separately, whereas fractions 1 and 2 were pooled together. The percentages of these fractions in the eluate estimated from the peak areas were practically identical.
Thus, according to the data of electrophoresis, immunoblotting, and gel-permeation chromatography the rhMIS preparation isolated using immunoaffinity chromatography contains, in addition to the homodimer, high molecular weight aggregates that under reducing conditions are dissociated to form a single fragment with molecular mass of 70 kD. Aggregation of both native MIS and rhMIS was also reported in some papers [19, 20]. So, it was important to estimate the biological activity of both the initial rhMIS preparation and its separate fractions.
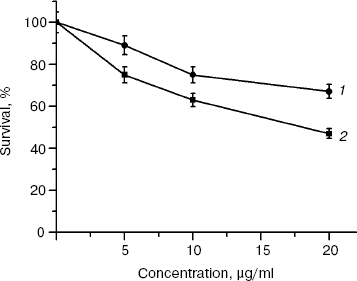
Biological activity of rhMIS. To determine a biological activity of the rhMIS preparation and its fractions with different molecular masses, we examined the ability of the proteins to influence survival (evaluated by the MTT-test) of human ovarian carcinoma line OVCAR8 cells and murine ovarian carcinoma line MOVCAR7 cells transfected with MISR II cDNA. The cells of these lines were incubated with rhMIS for 120 h. The data shown in Fig. 2 demonstrate that the rhMIS preparation decreased survival of both cell lines, and the inhibitory effect in the studied concentration range was dose-dependent. Notice that the effect of rhMIS is specific, because the tested preparation had no effect on survival of control SKOV3 cells in which the signal transmission from MIS receptors is known to be impaired (data on survivability of SKOV3 cells are shown further in Fig. 7). The MOVCAR7 cells were the most sensitive to rhMIS.
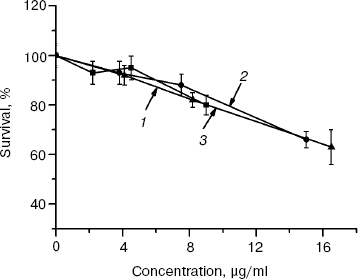
Biological activity of rhMIS fractions separated by gel filtration (140-kD homodimer and 280- and ~1000-kD oligomers) was tested by the effect of these fractions on survival of murine ovarian carcinoma (line MOVCAR7) cells (Fig. 3). The data show that all fractions possessed equal antiproliferative activity (calculated per mg protein) against the MOVCAR7 cells. When the activity is recalculated per mole protein, the biological activity of oligomeric proteins is significantly higher than that of the dimer. Thus, none of the fractions contained MIS inhibitors, which were reported to be present by some authors [21, 22] and required their elimination by additional purification steps.Fig. 2. Effect of rhMIS on survival of human OVCAR8 (1) and murine MOVCAR7 (2) ovarian carcinoma cells.
The data show that the isolated rhMIS protein, despite heterogeneity in molecular mass, possesses high biological activity against the model tumor cells expressing MISR II receptors. This allows the use of rhMIS in biological experiments without its separation into individual fractions.Fig. 3. Effect of rhMIS homodimer (140 kD) (3) and its aggregates (1000 (1) and 280 kD (2)) on survival of murine ovarian carcinoma MOVCAR7 cells (72 h incubation).
Effect of rhMIS on cell survival may depend on the number of its receptors on the cell membrane, adequate signal transmission upon rhMIS binding with these receptors, and total amount of MIS-binding proteins in the cell or the level of endocytosis of this protein. The results of evaluation of these parameters are given below.
Examination of binding of rhMIS with plasma membrane of tumor cells and its intracellular receptors and measurement of endocytosis rate by flow cytofluorometry. To characterize the expression level of rhMIS receptors in tumor cells and the possibility of receptor-mediated endocytosis of rhMIS, we examined the binding at 4°C and accumulation at 37°C of FITC-labeled rhMIS by various cell lines using flow cytofluorometry. It was demonstrated in preliminary experiments that the introduction of two FITC molecules per rhMIS molecule did not affect its biological activity and immunochemical properties (data not shown).
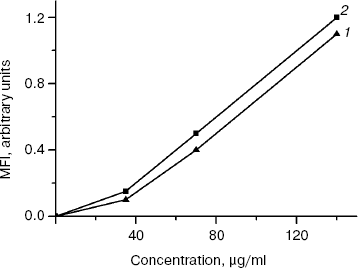
The binding of rhMIS with the MOVCAR7 and OVCAR8 cells was not high in the concentration range of 40-140 µg/ml and was proportional to its concentration. However, the curves of rhMIS binding did not have a saturation plateau even at maximal tested concentration of 140 µg/ml (1 µM of homodimer) (Fig. 4). The data indicate that rhMIS binding with surface receptors is of low affinity. At the same time, besides homodimer, about 30% tetramer and 30% oligomer of rhMIS are present in the preparation, so the actual molar concentration of rhMIS is lower than 1 µM. Taking into consideration that rhMIS has its biological effect at concentrations below 1 µM (Figs. 2 and 3), and increase in ligand concentration may lead to decrease in specificity of its binding with a receptor, we did not study the binding at higher concentrations of FITC-labeled rhMIS.
The level of rhMIS binding with MOVCAR7 cells was higher than with OVCAR8 cells, which is determined by the higher level of expression of rhMIS receptors on the membrane of murine ovarian carcinoma cells transfected with MISR II cDNA and correlates with higher sensitivity of these cells to rhMIS (Fig. 2).Fig. 4. Binding of FITC-labeled rhMIS with human ovarian carcinoma OVCAR8 cells (1) and murine ovarian carcinoma MOVCAR7 cells (2) depending on its concentration. The data of one of three typical experiments are presented. MFI, mean fluorescence intensity.
Experiments on binding of FITC-labeled rhMIS at 4°C with cultured in vitro human tumor cells of various lines, such as ovarian carcinoma SKOV3 and SKVLB, prostate carcinoma LNCaP and Du145, melanoma MS, and primary melanomas Mel-5, Mel-6, and Mel-8, showed that rhMIS binds with the surface of cell membrane of various tumor cells, but the binding level is low, thus suggesting low expression level of the receptors of this protein in membranes of these cells (table). The highest binding level was found in prostate cancer LNCaP cells and melanoma Mel-6.
Level of MIS-FITC (70 µg/ml) binding with plasma membranes of
tumor cells of various lines at 4°C, its accumulation at
37°C, endocytosis, and intracellular binding
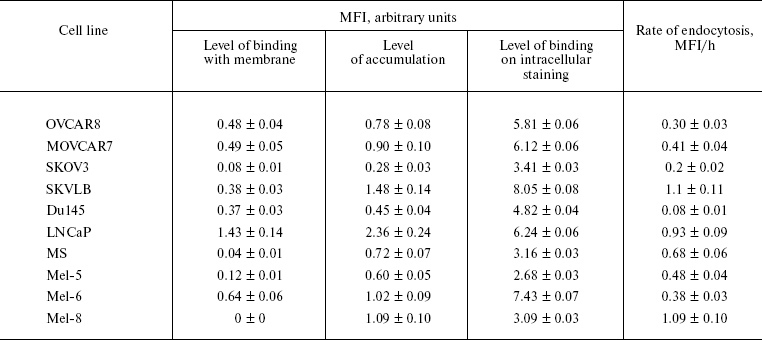
To measure the rate of rhMIS accumulation in various cell lines, the cells were incubated with FITC-labeled rhMIS at 37°C. The data presented in the table show that the mean fluorescence intensity (MFI) of cells after incubation with rhMIS-FITC at 37°C compared with 4°C is significantly higher, thus suggesting active endocytosis of rhMIS protein by tumor cells.
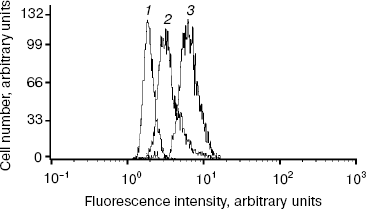
For study of intracellular binding of FITC-labeled rhMIS, we preliminarily examined specificity of the binding of this protein with intracellular receptors. To do this, the cells of studied lines were permeabilized followed by incubation with 20-fold excess of non-labeled rhMIS for 1 h before addition of FITC-labeled rhMIS. MFI of Du145 cells stained with FITC-labeled rhMIS after incubation with 20-fold excess of non-labeled protein and without it was 1.79 and 4.82 arbitrary units, respectively (Fig. 5), which indicates specificity of rhMIS binding with its intracellular receptors of no less than 60%. Analogous data supporting the specificity of rhMIS binding with the intracellular receptors were obtained in the case of other cell lines. The data on the intracellular binding of FITC-labeled rhMIS with the cells of various lines, which reflects the total amount of MIS binding proteins in cells, are presented in the table.
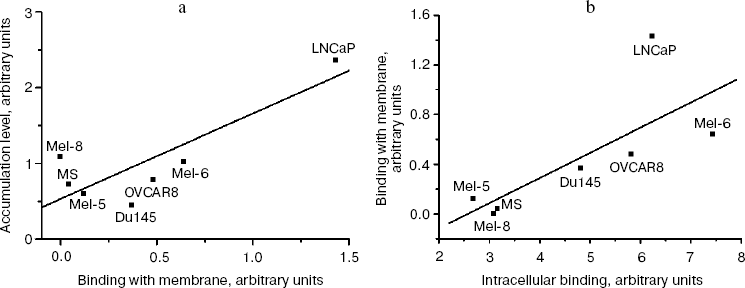
The highest rate of endocytosis of rhMIS was found in the cells of lines SKVLB, LNCaP, and Mel-8, the highest binding level in LNCaP and Mel-6, and the highest content of rhMIS-binding proteins in cells in LNCaP, SKVLB, and Mel-6. Analysis of correlation between the accumulation level of rhMIS-FITC in cells and its binding with the plasma membrane showed (Fig. 6a) that the accumulation of rhMIS-FITC in cells is in direct proportion with the level of its binding with the membrane (r = 0.83, p = 0.02, y = -0.21 + 0.19x), thus suggesting receptor-mediated endocytosis of MIS by tumor cells. The level of MIS binding with the membrane was in direct proportion to the intracellular level of MIS-binding proteins in tumor cells (r = 0.74, p = 0.05, y = -0.52 + 0.20x) (Fig. 6b).Fig. 5. Histogram of binding of FITC-labeled rhMIS (70 µg/ml) on the intracellular staining of human prostate Du145 line cells: 1) autofluorescence of the cells; 2) fluorescence of cells stained with rhMIS-FITC after incubation with 20-fold excess of the non-labeled protein; 3) fluorescence of cells stained with rhMIS-FITC.
Thus, we found that the major part of rhMIS receptors is localized inside of the cells, with less than 10% of MIS-binding proteins on the cell surface. The expression level of membrane-associated MIS receptors is therefore proportional to the level of this protein in the cell and determines the level of MIS accumulation upon its endocytosis.Fig. 6. Analysis of correlation between accumulation of MIS-FITC in cells and the level of its binding with the plasma membrane of the cells (a) and of correlation between the level of MIS-FITC binding with the membrane and the level of its binding with intracellular receptors (b).
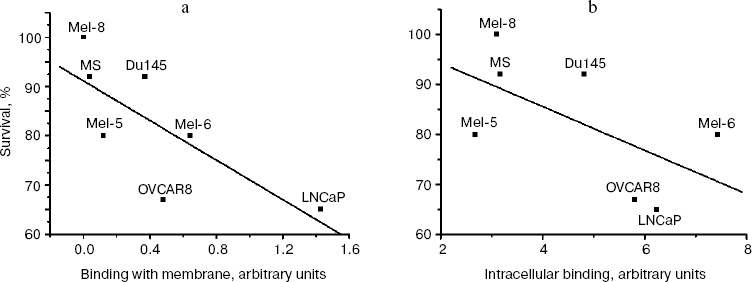
Relationship between level of MIS receptor expression and biological response of various tumor line cells to rhMIS. Examination of biological activity of rhMIS against various tumor cell lines demonstrated that survivability of the cells decreased in the presence of rhMIS with highest antiproliferative activity against LNCaP cells (Fig. 7). The rhMIS had no effect on survival of SKOV3 and SKVLB cells, which supports the literature data and is known to be associated with the defect in the system of signal transmission from MISR II in these cells [16]. A comparison of survivability of these cells with the level of rhMIS-FITC binding with the plasma membranes of human tumor cells (lines SKOV3 and SKVLB were excluded from analysis) revealed a statistically significant inverse correlation between cell survival in presence of rhMIS (20 µg/ml) and level of rhMIS binding on the cell membrane (r = -0.75, p = 0.04, y = 91.13 - 20.11x) (Fig. 8a). At the same time, survival of the cells also appears to be inversely proportional to the total level of MIS-binding proteins determined by intracellular staining (Fig. 8b), but this dependence was less statistically significant (r = -0.60, p = 0.15, y = 102.95 - 4.35x). We found no correlation between cell survival and the levels of accumulation and endocytosis of MIS (data not shown).
Fig. 7. Survival of human tumor cells of different lines after 120 h of incubation with rhMIS (20 µg/ml).
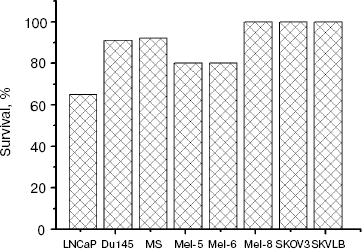
Fig. 8. Correlation between human tumor cell survival in the presence of rhMIS and the level of MIS-FITC binding with the plasma membrane (a) and with intracellular receptors (b).
DISCUSSION
In this study we have shown that the rhMIS preparation isolated from the culture medium of CHO cells transfected with cDNA of this protein contains (after immunoaffinity purification) not only homodimer, but also oligomeric rhMIS complexes. Analogous results were obtained when native MIS was isolated from bovine testicles [19]. According to the literature, some cytokines, such as adiponectin, are active only in the form of oligomeric complexes. In particular, it had been shown that only after oligomerization into hexamers and higher order complexes adiponectin can activate the NF-kappaB transcription factor, which is necessary for expression of at least some of its biological effects [23]. The testing of biological activity of rhMIS fractions separated by gel filtration has shown that all rhMIS fractions, independently on their aggregation degree, decrease survival of tumor cells, that is, they possess biological activity. An explanation might be that either MIS is active in oligomeric form or its biological activity is only determined by the C-terminal fragment, which is cleaved off upon limited proteolysis of this protein [21]. It is worth noting that the amount of produced C-terminal fragment may not depend on MIS aggregation degree or may be even higher when MIS is present in the form of oligomers.
Our experiments have shown that the expression level of receptors on the plasma membrane is low in comparison with the level of intracellular receptors, and rhMIS binding with surface receptors is of low affinity and does not reach saturation in the rhMIS concentration range up to 1 µM. It is possible that oligomeric rather than monomeric MIS binds with MISRII, as in the case of adiponectin, and then the effective concentration of this protein is different. At the same time, the MIS concentration used in experiments on binding quite corresponds to its biological activity, so the use of higher concentrations is inappropriate.
It was demonstrated by the example of another protein of this family, TGFbeta. that after internalization of the TGFbeta-receptor complex an activation of I type receptor and actuation of signaling cascade occur, and recycling of TGFbeta receptors in endosomes and vesicles is activated [22, 24]. The high intracellular MIS receptor level we found might be important for this process. This supposition is supported by other data. In particular, it has been found that the number of MIS receptors exposed at the surface of plasma membrane varied between the cells of tested lines and was proportional both to the total amount of MIS-binding proteins and to the level of rhMIS-FITC accumulation upon its endocytosis. The found statistically significant correlation between the level of rhMIS-FITC binding with cellular membrane and survival of tumor cells in the presence of rhMIS and the absence of statistically significant dependence of cell survival on the level of intracellular MIS-binding proteins suggest that it is the membrane-associated MIS receptors that determine its biological effect against tumor cells.
REFERENCES
1.Coughlin, J. P., Donahoe, P. K., Budzik, G. P., and
MacLaughlin, D. T. (1987) Mol. Cell Endocrinol., 49,
75-86.
2.Visser, J. A., Olaso, R., Verhoef-Post, M., Kramer,
P., Themmen, A. P., and Ingraham, H. A. (2001) Mol. Endocrinol.,
15, 936-945.
3.Visser, J. A. (2003) Mol. Cell Endocrinol.,
211, 65-73.
4.Clarke, T. R., Hoshiya, Y., Yi, S. E., Liy, X.,
Lyons, K. M., and Donahoe, P. K. (2001) Mol. Endocrinol.,
15, 946-959.
5.Gouedard, L., Chen, Y. G., Thevenet, L., Racine,
C., Borie, S., Lamarre, I., Josso, N., Massague, J., and di Clemente,
N. (2000) J. Biol. Chem., 275, 27973-27978.
6.Visser, J. A., Olaso, R., Verhoef-Post, M., Kramer,
P., Themmen, A. P., and Ingraham, H. A. (2001) Endocrinology,
131, 1481-1488.
7.Jamin, S. P., Arango, N. A., Mishina, Y., Hanks, M.
C., and Behringer, R. R. (2002) Nat. Genet., 32,
408-410.
8.Jamin, S. P., Arango, N. A., Mishina, Y., Hanks, M.
C., and Behringer, R. R. (2003) Mol. Cell Endocrinol.,
211, 15-19.
9.Renaud, E. J., MacLaughlin, D. T., Oliva, E.,
Rueda, B. R., and Donahoe, P. K. (2005) Proc. Natl. Acad. Sci.
USA, 102, 111-116.
10.Masiakos, P. T., MacLaughlin, D. T., Maheswaran,
S., Teixeira, J., Fuller, A. F., Jr., Shah, P. C., Kehas, D. J.,
Kenneally, M. K., Dombkowski, D. M., Ha, T. U., Preffer, F. I., and
Donahoe, P. K. (1999) Clin. Cancer Res., 5,
3488-3499.
11.Barbie, T. U., Barbie, D. A., MacLaughlin, D. T.,
Maheswaran, S., and Donahoe, P. K. (2003) Proc. Natl. Acad. Sci.
USA, 100, 15601-15606.
12.Connolly, D. C., Bao, R., Nikitin, A. Y.,
Stephens, K. C., Poole, T. W., Hua, X., Harris, S. S., Vanderhyden, B.
C., and Hamilton, T. C. (2003) Cancer Res., 63,
1389-1397.
13.Donahoe, P. K., Swann, D. A., Hayashi, A., and
Sullivan, M. D. (1979) Science, 205, 913-915.
14.Roberts, L. M., Hirokawa, Y., Nachtigal, M. W.,
and Ingraham, H. A. (1999) Dev. Biol., 208, 110-112.
15.Parry, R. L., Chin, T., Epstein, J., Hudson, P.
L., Powell, D. M., and Donahoe, P. K. (1992) Cancer Res.,
52, 1182-1186.
16.Wallen, J. W., Cate, R. L., Kiefer, D. M.,
Riemen, M. W., Martinez, D., Hoffman, R. M., von Hoff, D. D., Donahoe,
P. K., Pepinsky, R. B., and Oliff, A. (1989) Cancer Res.,
49, 2005-2011.
17.Bradley, G., Naik, M., and Ling, V. (1989)
Cancer Res., 49, 2790-2796.
18.Mosmann, T. (1983) J. Immunol. Meth.,
65, 55-63.
19.Picard, J. Y., and Josso, N. (1984) Mol. Cell
Endocrinol., 34, 23-29.
20.Shima, H., Donahoe, P. K., Budzik, G. P.,
Kamagata, S., Hudson, P., and Mudgett-Hunter, M. (1984)
Hybridoma, 3, 201-204.
21.Szotek, P. P., Pieretti-Vanmarcke, R., Masiakos,
P. T., Dinulescu, D. M., Connolly, D., Foster, R., Dombkowski, D. M.,
Preffer, F. I., MacLaughlin, D. T., and Donahoe, P. K. (2006) Proc.
Natl. Acad. Sci. USA, 103, 11154-11159.
22.Penheiter, S. G., Mitchell, H., Garamszegi, N.,
Edens, M., Dore, J. E., Leof, E., Jr., and Leof, E. (2002) Mol.
Cell. Biol., 22, 4750-4759.
23.Tsao, T. S., Murrey, H. E., Hug, C., et al.
(2002) J. Biol. Chem., 277, 29359-29362.
24.Ito, T., Williams, J. D., Fraser, D. J., and
Phillips, A. O. (2004) J. Biol. Chem., 279,
25326-25332.