Vectorial Charge Transfer Reactions on the Donor Side of Manganese-Depleted and Reconstituted Photosystem 2 Core Complexes
I. O. Petrova, V. N. Kurashov, A. A. Zaspa, A. Yu. Semenov, and M. D. Mamedov*
Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, 119234 Moscow, Russia; fax: (495) 939-3181; E-mail: mahirmamedov@yandex.ru; mamedov@genebee.msu.ru* To whom correspondence should be addressed.
Received December 18, 2012
The light-induced functioning of photosystem 2 (PS 2) is directly linked to the translocation of both electrons and protons across the membrane, which results in the formation of transmembrane electric potential difference (ΔΨ). Generation of ΔΨ due to S-state transitions of the water oxidation complex was demonstrated for the first time in Mn-depleted and reconstituted PS 2 core complexes incorporated into liposomes. The kinetics and relative amplitudes of the electrogenic reactions in dark-adapted samples during S1→S2, S2→S3, and S4→S0 transitions in response to the first, second and third laser flashes were comparable to those obtained in the intact PS 2 core particles. These results expand current understanding of the nature and mechanisms of electrogenic (vectorial) reactions due to a charge transfer on the donor side of PS 2.
KEY WORDS: photosystem 2, apo-WOC-PS 2, photoactivation, S-state transitions, proteoliposomes, direct electrometric techniqueDOI: 10.1134/S0006297913040081
Abbreviations: PS 2, photosystem 2; P680, primary electron donor; QA and QB, primary and secondary quinone acceptors, respectively; RC, reaction center; WOC, water oxidation complex; YZ and YZ•, reduced and neutral radical form of Tyr161 residue of D1 protein, respectively; ΔΨ, transmembrane electric potential difference.
The pigment–protein complex of photosystem 2 (PS 2) is located in
thylakoid membranes of cyanobacteria, algae, and plants, and it
catalyzes the light-driven oxidation of water to molecular oxygen and
reduction of plastoquinone to plastohydroquinone. The structures of
dimeric PS 2 core complexes from thermophilic cyanobacteria have been
solved by X-ray crystallography with atomic resolution [1-3]. Each monomer of the enzyme
with molecular mass of ~350 kDa consists of 20 protein subunits
(17 integral and three peripheral) and ~70 cofactors. In so doing, all
the organic and inorganic cofactors involved in charge separation
reactions are located on the central D1 and D2 subunits that form the
reaction center (RC) heterodimer.
Functionally, PS 2 can be described in terms of three structural domains: central photochemical, plastoquinone reducing, and water-oxidizing. The water-oxidizing domain or water oxidation complex (WOC) in thylakoids is located on the lumenal (donor) side of PS 2. It contains four Mn ions, one Ca2+ ion (Mn4Ca cluster), surrounding amino acid ligands, and water molecules [3].
Excitation energy of light absorbed by CP43 and CP47 integral antenna pigments in the picosecond time range is transferred to the primary electron donor P680 composed of a dimer of chlorophylls. Excitation of P680 results in formation of the charge separated state between primary electron donor – a dimer of chlorophyll P680 and primary quinone acceptor QA (P680+QA−) with participation a monomeric chlorophyll and pheophytin in the active branch A. A very high potential oxidant, P680+, oxidizes the redox-active tyrosine residue YZ (Tyr161 of the D1 protein) forming the neutral radical YZ•. This radical, in turn, oxidizes the Mn cluster, while QA– reduces the secondary plastoquinone QB. Subsequent charge separations result in further oxidation of the Mn cluster and reduction of twice-reduced QB (QB2−) to plastoquinol (QBH2). During each catalytic cycle, the Mn cluster cycles via five kinetically characterized intermediate states labeled Si (i = 0-4) [4-8]. With four sequential oxidations from S0 to S4, the water-oxidizing complex (WOC) is capable of splitting water into dioxygen, four electrons, and four protons. Four protons are transferred from the Mn complex or its ligand environment to the bulk phase with the release pattern of 1 : 0 : 1 : 2 for S0→S1→S2→S3→S0 transitions, while an electron is released in every single transition [9-14].
It should be noted that water oxidation and O2 evolution are completely inhibited upon depletion of Mn ions from PS 2. The Mn-depleted complexes are usually designated as apo-WOC-PS 2. The WOC function can be assembled in the presence of the free inorganic ions (Mn2+, Ca2+, Cl−) and PS 2, which are capable of performing this complex process ([15, 16] and references therein). This light-dependent process, which also occurs upon assembling of a manganese cluster in vivo [16, 18], is called photoactivation.
Although the internal electron and proton transfer processes form a complicated overall picture of the PS 2 complex function, this function can be studied by measurement of the transmembrane electric potential difference (ΔΨ) using a direct electrometric technique ([19, 20] and references therein) and an electrochromic shift of the absorption bands of carotenoids [19]. Earlier results in liposome-reconstituted, oxygen-evolving PS 2 core particles [19-21], as well as in thylakoids from higher plants [19], indicate the electrogenic reactions corresponding to the S-state transitions of the WOC on the first three laser flashes.
In the present work, the direct electrometric technique was applied for the first time to study the electrogenesis associated with reactions of electron and proton transfer during S-transitions in Mn-reconstituted (photoactivated) apo-WOC-PS 2 core particles incorporated into liposomes. The direct electrometry is an exclusively sensitive technique that allows registering kinetics of intraprotein charge transfer over distance >0.5 Å in the direction perpendicular to the plane of a membrane and estimating the relative contribution of individual electrogenic reactions (dielectrically weighted transmembrane distances between cofactors) to overall ΔΨ upon a single turnover of the photosynthetic RC [22].
MATERIALS AND METHODS
Oxygen-evolving PS 2 core complexes were isolated from spinach as described in [23] with minor modifications. To extract Mn, Ca, and extrinsic proteins, PS 2 core complexes were treated with 0.8 M Tris (pH 8.3) buffer for 30 min [24].
Apo-WOC-PS 2 core complexes were reconstituted into phospholipid (L-α-phosphatidylcholine, type II-S; Sigma, USA) vesicles as described in [25]. Phospholipid suspension in 50 mM HEPES-HCl buffer (pH 7.5) containing 0.8% (w/v) n-octyl-β-D-glucopyranoside (Sigma) was sonicated until clarity. The PS 2 core particles were incorporated into the liposomes by mixing a small volume of sample with liposome suspension (standard lipid-to-protein ratio of 30 (w/w)) and incubation of the mixture for 30 min at 4ºC in the dark. After mixing, the samples were loaded on a Sephadex G50 column (NAP5; Pharmacia, Sweden) previously equilibrated with 25 mM HEPES-NaOH (pH 7.5) buffer containing 5 mM CaCl2. The proteoliposomes were collected employing stepwise elution with the same solution.
Light-induced reassembly of functional WOC in apo-WOC-PS 2 core particles was achieved as follows: the proteoliposomes in assay medium (300 mM sucrose, 15 mM NaCl, 10 mM CaCl2, 10 μM MnCl2, 25 mM MES-NaOH, pH 6.5) were incubated for 3 min by gentle stirring under weak continuous light (20 mV/cm2). Potassium ferricyanide (1 mM) as an electron acceptor for QA− was trapped inside the vesicles. The rate of oxygen evolution (~1500 μmol O2/h per mg Chl) for proteoliposomes containing intact PS 2 core particles corresponded to 100% activity. The oxygen-evolving activities of apo-WOC-PS 2 samples before and after photoactivation were 30-50 and ~830-850 μmol O2/h per mg Chl, respectively.
Note that in PS 2 membrane fragments the efficiency of photoactivation also depends on bicarbonate ([15, 16] and references therein).
The rate of steady-state O2 evolution was measured with a Clark-type O2 electrode (Hansatech, UK). Chlorophyll concentration during the measurements was 10 μg/ml.
Voltage transients were measured using the direct electrometric technique [22]. In the measuring system, Ag/AgCl electrodes record the voltage between the two compartments of a cell separated by a measuring membrane consisting of a lipid-impregnated collodion film. The association of proteoliposomes with the artificial lipid membrane is achieved by addition of MgCl2 to the assay medium for neutralization of negative charges on the membrane surface. A Quantel Nd-YAG laser (France) (emission wavelength, 532 nm; full width at half maximum, 12 ns) was used as the source of light pulses.
Kinetic signals were analyzed using program packages Pluk [26] and Origin (OriginLab Corporation, USA).
RESULTS AND DISCUSSION
Figure 1 shows electrometrically detected voltage transients induced by the 1st laser flash in dark-adapted liposome-reconstituted Mn-depleted and Mn-reconstituted (+Mn) core particles. The negative sign of ΔΨ indicates that the donor side of the enzyme is located at the external surface of the proteoliposomal membrane [21, 24]. The lack of membrane impermeable sodium dithionite effect on the amplitude of the voltage transient indicates a highly asymmetric orientation of PS 2 core complexes in the liposomes (~90% of the complexes are facing donor side outside) [25]. The maximum amplitude fast phase (the rise time is shorter than the time resolution of the experimental setup (~100 ns)) corresponds to charge separation in PS 2 RC between P680 and QA and subsequent P680+ reduction by electron transfer from redox active tyrosine YZ (YZ•QA−) [22]. The lack of additional electrogenicity in the kinetics of the voltage transient in proteoliposomes in the sub-millisecond time range indicates that the rate of electron transfer from YZ to P680+ is not significantly slowed after depletion of manganese.
Fig. 1. Voltage transients induced by the 1st laser flash in proteoliposomes containing dark-adapted apo-WOC-PS 2 (–Mn) and Mn-reconstituted (+Mn) core complexes. The kinetic traces were normalized by the amplitude of the fast electrogenic phase observed in apo-WOC-PS 2 (–Mn) complexes and ascribed to formation of YZ•QA–. The apo-WOC-PS-2 cores were suspended in the assay medium containing 300 mM sucrose, 10 mM CaCl2, 15 mM NaCl, and 25 mM MES/NaOH at pH 6.5. For the photoactivation of the samples, the medium was supplemented with 10 µM MnCl2. K3[Fe(CN)6] was used as an electron acceptor at 1 mM (see “Materials and Methods”). The inset shows the difference of voltage transients between Mn-reconstituted and apo-WOC-PS 2 samples. All experiments were performed at room temperature (22 ± 1ºC). Here and in the other figures, the vertical arrows indicate the time of the laser flash.
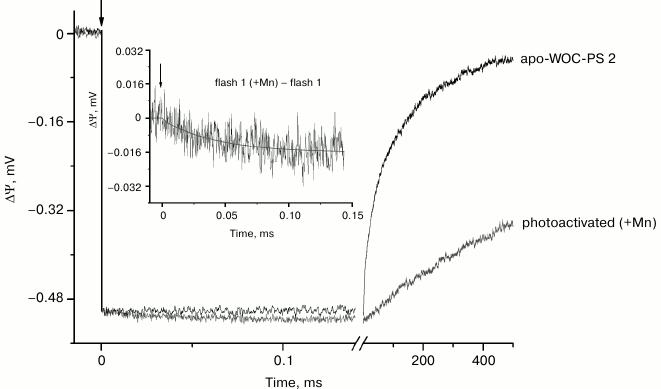
In oxygen-evolving PS 2, photoreduced QA is reoxidized by a secondary quinone acceptor QB during 0.1-0.3 ms [5]. However, the electron transfer reaction between QA and QB is not functional in PS 2 associated with a phospholipid-impregnated collodion film. This is due to extraction of loosely bound QB into artificial lipid membrane [22]. Recombination of the electron between QA− and the neutral radical YZ• occurs in PS 2 complexes depleted of Mn4Ca cluster and three extrinsic proteins [27-29]. The kinetics of ΔΨ decay in these preparations can be approximated by two exponentials with characteristic times of ~30 ms (relative amplitude ~43%) and ~240 ms (~57%). These components reflect sub-states of the enzyme and are probably related to charge recombination between QA− and YZ• [28, 30].
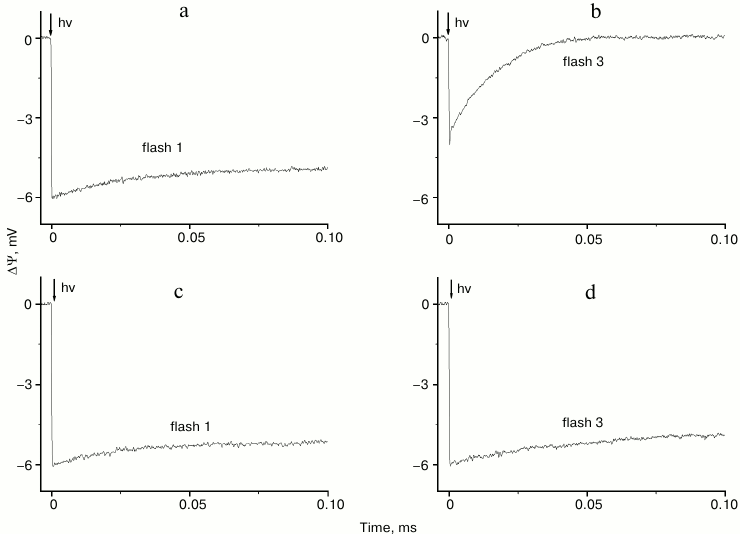
Thus, in apo-WOC-PS 2 core complexes reconstituted into liposomes the first flash apparently yielded YZ•QA− (Fig. 2a), while the stable charge separation induced by following flashes was largely prevented because QA− was still present (Fig. 2b). The rapid decay of the voltage transient under these conditions corresponds to electron recombination from pheophytin to P680+. However, when potassium ferricyanide (K3[Fe(CN)6]) was added as an artificial electron acceptor, QA− decayed rapidly between flashes (in <0.5 s), and electron transfer between YZ and QA occurred in response to subsequent flashes (Fig. 2, c and d). Under these conditions, proton uptake on the acceptor side of PS 2 did not occur [25], and it produces the highest yield of the photoassembly ([31] and references therein).
Fig. 2. Voltage transients on flashes 1 and 3 in dark-adapted, liposome-reconstituted apo-WOC-PS 2 core particles in the absence (a, b) and presence of 1 mM K3[Fe(CN)6] (c, d).
In Mn-depleted PS 2 core complexes the oxygen-evolving activity was negligible, while in the presence of exogenously added Mn2+ the relative rate of oxygen evolution under steady state illumination was ~55% of that observed for control samples (see “Materials and Methods” and [32]).
It should be noted that comparative analysis of data derived from measurement of chlorophyll fluorescence induction kinetics of Mn-reconstituted apo-WOC-PS 2 core complexes in solution or reconstituted into liposomes suggests that the lipid environment stimulates the steady-state rate of electron transfer from the manganese cluster to plastoquinone [32]. The effect of lipids is likely based on keeping PS 2 RC in its optimal structure for efficient functioning.
After dark adaptation of liposome-reconstituted intact PS 2 core particles, all WOC were synchronized mainly in the first oxidized state S1 [18, 19] and the O2 yield was observed on third and seventh flashes [19]. As to dark-adapted Mn-reconstituted apo-WOC-PS 2 samples, O2 oscillation pattern showed a significant O2 yield on the 2nd flash, peaking on the 3rd and 4th flashes (not shown). In so doing, no oscillations were observed beyond the 6th flash (see also [31] and references therein). However, we cannot unambiguously apply the results concerning the rate of oxygen evolution in response to a series of light flashes (flash lifetime ~60 µs) of Mn-reconstituted apo-WOC-PS 2 complexes to data obtained in the present work. In the latter case, laser pulses were used as the source of light (see “Materials and Methods”).
As seen from Fig. 1, addition of Mn to apo-WOC-PS 2 samples upon photoactivation results in significant slowing of the decay of ΔΨ due to prevention of charge recombination and appearance of the small additional electrogenic phase in the sub-millisecond time range indicating effective interaction between manganese and YZ•.
Analysis of the difference between kinetic curves induced by a single flash in the presence and absence of Mn reveals that the electrogenicity is related to the reduction of YZ• by Mn. This difference [1f(Mn) – 1f] presented in the inset to Fig. 1 is characterized by lifetime (τ) of ~40 µs and relative amplitude ~3% of the amplitude of the phase ascribed to YZ•QA− formation. The value of τ is close to the rate of oxidation of a single Mn2+ ion by YZ• ([33] and references therein) corresponding to the S1→S2 transition of the WOC in intact PS 2 core complexes [19, 20]. It should be noted that there is no proton release on the S1→S2 transition, so a positive charge is accumulated on the donor side [8, 13, 14, 19]. Thus, vectorial electron transfer from Mn to YZ• on the first laser flash can be presumed to correspond to the S1→S2 transition in Mn-reconstituted apo-WOC-PS 2 samples.
The voltage transients induced by the 2nd (S2→S3 transition) and 5th laser flashes in dark-adapted samples after normalization of the fast amplitudes corresponding to the formation of YZ•QA− are shown in Fig. 3a. The 5th flash mainly induced the S1→S2 transition. It is similar to the 1st flash, but without possible contribution from electron transfer to the non-heme iron located between quinones [19, 25]. Therefore, the difference between kinetic traces induced by the 2nd and 5th flashes (Fig. 3b) reflects the difference between the 2nd and 1st flashes. The photoelectric responses induced by the 1st flash are due to electron transfer from YZ to QA and subsequent re-reduction of YZ• by Mn. Thus, subtraction of the 1st flash-induced kinetic trace from kinetic traces induced by subsequent flashes eliminates contributions derived from Mn→YZ and YZ→QA electron transfer reactions. It is therefore reasonable to assume that the electrogenicity observed during S2→S3 and the following S3→S4→S0 transitions related to the turnover of the WOC could be attributed to proton transfer from the substrate water molecule [14] or amino acids in the vicinity of the Mn complex to the bulk phase [8, 19]. Analysis of kinetics induced by the 2nd and 5th laser flashes (Fig. 3b) shows that the exponential phase has a lifetime of ~220 µs, whereas its amplitude is ~6.8% of the amplitude of the fast phase corresponding to the YZ•QA− formation. It should be noted that the 2nd and 5th kinetic curves do not clearly differ until ~0.1 ms, while the traces induced by the 1st flash in the presence and absence of Mn2+ (Fig. 1) become distinguishable after ~0.05 ms.
Fig. 3. Voltage transients induced by the 2nd and 5th laser flashes (a) and their difference (b) in dark-adapted Mn-reconstituted PS 2 core particles incorporated into liposomes. The kinetic traces here and in Fig. 4 were normalized by the amplitude of the 5th flash-induced fast phase due to YZ•QA– formation. Flash spacing was 0.5 s. The difference of voltage transients induced by the 2nd and 5th laser flashes after illumination by nine laser flashes (c).
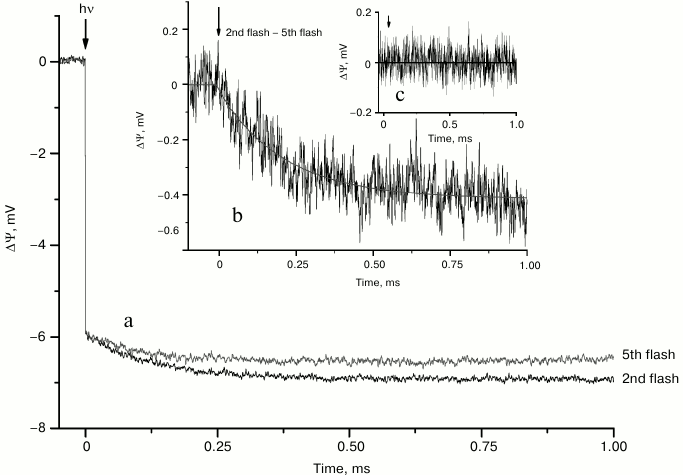
The difference of voltage transients induced by the 2nd and 5th laser flashes registered after a series from nine laser flashes was close to zero (Fig. 3c). This result demonstrates that in previously excited Mn-reconstituted apo-WOC-PS 2 samples all S-states are presented equally, while in dark-adapted samples the S1 state probably prevails.
An additional electrogenic phase in the kinetics of the voltage transient induced by the 2nd flash (S2→S3 transition) at pH 6.5 in intact core particles and in thylakoids has τ ~270 and ~300 μs, respectively [19]. This phase is ascribed to a proton transfer into the aqueous phase due to pK shift and deprotonation of an unidentified amino acid in the vicinity of the Mn cluster [19]. We thus assume that an additional ~220 µs electrogenic phase in the kinetics of the voltage transient of the Mn-reconstituted apo-WOC-PS 2 on the S2→S3 transition is due to proton transfer from the Mn complex into the aqueous bulk phase.
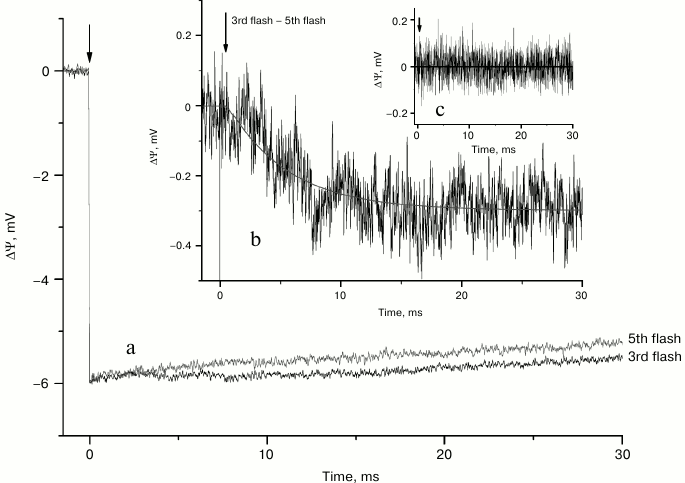
Generation of ΔΨ induced by the 3rd and 5th laser flashes is shown in Fig. 4a. The difference of voltage transients (3f – 5f) (Fig. 4b) revealed much slower additional rising of electrogenic component with a characteristic time of ~5 ms and relative extent of ~5% of the formation of YZ•QA−. In so doing, the difference of voltage transients induced by the 3rd and 5th flashes and registered after nine laser flashes (Fig. 4c) as in the case of the S2→S3 transition (Fig. 3c) was about zero.
Fig. 4. Voltage transients induced by the 3rd and 5th laser flashes (a) and their difference (b) in dark-adapted liposome-reconstituted Mn-reconstituted PS 2 core particles. Difference of voltage transients induced by the 3rd and 5th laser flashes after illumination by nine laser flashes (c).
In intact PS 2 complexes, the central event in S4 formation seems to be deprotonation of a residue near the Mn complex with a lifetime of ~200 µs, while the final S4→S0 transition of the cycle is associated with the release of oxygen and another proton [8].
As can be seen from the difference of photoelectric responses on the 3rd and 5th laser flashes (3f – 5f), the voltage kinetics lack a sub-millisecond component but reveal a ~5 ms component (Fig. 4b). This finding most probably indicates that the proton release accompanying the S3→S4 transition is electrically silent, while proton release during S4→S0 transition is electrogenic. It should be noted that the lifetime of the S4→S0 transition was estimated to be ~1.2 ms in thylakoids [19] and PS 2 membrane fragments [13] but ~4.6-6 ms in intact PS 2 core complexes [19, 20].
Similar values of relative amplitudes of the electrogenic phases with significantly different kinetics of proton transfer reactions during the S2→S3 and S4→S0 transitions of Mn-reconstituted apo-WOC-PS 2 samples imply that protons probably cross approximately similar dielectrically weighted distances, but apparently by different pathways.
The S0→S1 transition induced by the 4th flash most likely involves oxidation of the Mn complex by YZ• as well as charge-compensating deprotonation of an amino acid residue close to the manganese cluster [8, 13]. The subtraction of the 5th from the 4th flash-induced photoelectric signal does not reveal any difference (data not shown). Since the 5th flash is considered as similar to the 1st flash, the photoelectric signals in both cases are due to the charge separation between P680 and QA, reduction of P680+ by YZ, and subsequent re-reduction of YZ• by Mn. In addition, the 4th flash-induced S0→S1 transition includes proton release. The lack of an additional voltage induced by the 4th and 5th flashes indicates that proton release during S0→S1 is not electrogenic.
Thus, the results of the present work show that the kinetics and relative amplitudes of the electrogenic phases during the S-state transitions in Mn-reconstituted apo-WOC-PS 2 complexes are comparable to those obtained in intact PS 2 core complexes (see table). Almost the full reconstruction of electrogenic S1→S2, S2→S3, and S4→S0 transitions occurs even in the absence of the extrinsic proteins, and this implies that the S-states are stable for at least 0.5 s. However, a highly conserved extrinsic manganese-stabilizing protein has been suggested as essential for maximum yield of recovering of the oxygen evolution rate ([16, 34] and references therein). Indeed, the relative rate of oxygen evolution upon reassembling WOC in our experiments did not exceed ~55%. Thus, one can conclude that the full reconstruction of electrogenic reactions due to S-state transitions is insufficient for achieving the entire recovery of WOC function.
Relative amplitudes and lifetimes of electrogenic reactions of water
oxidation complex of photosystem 2
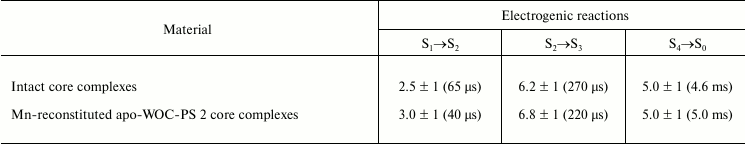
Note: Amplitudes of electrogenic reactions are given as a percentage of
the fast phase corresponding to
YZ•QA− formation
(100%). Characteristic times of reactions are presented in
parentheses.
In the present work, we applied for the first time a sensitive electrometric technique to study electrogenic reactions due to intraprotein vectorial electron and proton transfer during the catalytic cycle of water oxidation in manganese-depleted and reconstituted PS 2 core complexes. In dark-adapted samples, the electrogenic reactions observed in response to the 1st, 2nd, and 3rd laser flashes are presumably associated with the S1→S2 (electron transfer from Mn to YZ•), S2→S3, and S4→S0 (proton transfer in the opposite direction from Mn complex or its immediate environment into the aqueous bulk phase) transitions. The lack of additional voltage in the kinetics of the photoelectric response on the 4th laser flash indicates that proton release during the S0→S1 transition is not electrogenic. Thus, only two of the four proton transfer reactions during the catalytic cycle of water oxidation are electrogenic (S2→S3 and S4→S0 transitions). It should be noted that the electrogenicity due to electron transfer from Mn to YZ• (~3%) was also observed in the S0→S1, S2→S3, and S4→S0 transitions. These results expand current understanding of the nature and mechanisms of electrogenic reactions during the S-state transitions in PS 2 reaction centers and provide a necessary step in development of efficient systems of transformation of solar energy.
This work was supported financially by the Russian Foundation for Basic Research (grants 11-04-00818-a, 12-04-00821-a, 11-04-92503-a), by the DFG-RFBR Cooperation Project (grant No. 11-04-91330), by CRDF (RUB1-7029-MO-11), and by the Russian Ministry for Education and Science (16.512.12.2010).
REFERENCES
1.Ferreira, K. N., Iverson, T. M., Maghlaoui, K.,
Barber, J., and Iwata, S. (2004) Science, 303,
1831-1838.
2.Guskov, A., Kern, J., Gabdulkhakov, A., Broser, M.,
Zouni, A., and Saenger, W. (2009) Nat. Struct. Mol. Biol.,
16, 334-342.
3.Umena, Y., Kawakami, K., Shen, J.-R., and Kamiya,
N. (2011) Nature, 473, 55-60.
4.Kok, B., Forbush, B., and McGloin, M. (1970)
Photochem. Photobiol., 11, 457-475.
5.Shinkarev, V. P. (2004) in Photosystem II:
Oxygen Evolution and Chlorophyll a Fluorescence Induced by Multiple
Flashes (Papageorgiou, G. C., and Govindjee., eds.) Kluwer Academic
Publishers, Dordrecht, pp. 197-229.
6.Wydrzynski, T. J., and Satoh, K. (2005)
Photosystem II: The Light-Driven Water:Plastoquinone
Oxidoreductase, Springer, New York.
7.Renger, G., and Kuhn, P. (2007) Biochim.
Biophys. Acta, 1767, 458-471.
8.Dau, H., and Haumann, M. (2007) Biochim.
Biophys. Acta, 1767, 472-483.
9.Rappaport, F., and Lavergne, J. (1991)
Biochemistry, 30, 10004-10012.
10.Lavergne, J., and Junge, W. (1993) Photosynth.
Res., 38, 279-296.
11.Bogershausen, O., and Junge, W. (1995)
Biochim. Biophys. Acta, 1230, 177-185.
12.Schlodder, E., and Witt, H. T. (1999) J. Biol.
Chem., 274, 30387-30392.
13.Haumann, M., Liebisch, P., Muller, C., Barra, M.,
Grabolle, M., and Dau, H. (2005) Science, 310,
1019-1021.
14.Noguchi, T. (2008) Philos. Trans. R Soc.
B., 363, 1189-1195.
15.Baranov, S. V., Tyryshkin, A. M., Katz, D., and
Klimov, V. V. (2004) Biochemistry, 43, 2070-2079.
16.Dasgupta, J., Ananyev, G. M., Dismukes, G. C.,
Ananyev, G. M., and Klimov, V. V. (2008) Coord. Chem. Rev.,
252, 347-360.
17.Ono, T. A., and Mino, H. (1999)
Biochemistry, 38, 8778-8785.
18.Magnuson, A., Rova, M., Mamedov, F., Fredriksson,
P.-O., and Styring, S. (1999) Biochim. Biophys. Acta,
1411, 180-191.
19.Haumann, M., Mulkidjanian, A., and Junge, W.
(1997) Biochemistry, 36, 9304-9315.
20.Mamedov, M. D., Beshta, O. E., Gurovskaya, K. N.,
Mamedova, A. A., Neverov, K. D., Samuilov, V. D., and Semenov, A. Yu.
(1999) Biochemistry (Moscow), 64, 504-509.
21.Semenov, A., Cherepanov, D., and Mamedov, M.
(2008) Photosynth. Res., 98, 121-130.
22.Semenov, A. Yu., Mamedov, M. D., and Chamorovsky,
S. K. (2006) in Photosystem I. The Light-Driven
Plastocyanine:Ferredoxin Oxidoreductase (Golbeck, J. H., ed.)
Springer, Dordrecht, pp. 319-424.
23.Ghanotakis, D. F., Demetriou, D. M., and Yocum,
C. F. (1987) Biochim. Biophys. Acta, 891, 15-21.
24.Gopta, O. A., Tyunyatkina, A. A., Kurashov, V.
N., Semenov, A. Yu., and Mamedov, M. D. (2008) Eur. Biophys. J.,
37, 1045-1050.
25.Mamedov, M. D., Tyunyatkina, A. A., Siletsky, S.
A., and Semenov, A. Yu. (2006) Eur. Biophys. J., 35,
647-654.
26.Kalaidzidis, Ya. L., Gavrilov, A. V., Zaitsev, P.
V., Kalaidzidis, A. L., and Korolev, E. V. (1997) Programm. Computer
Software, 23, 206-212.
27.Nixon, P. J., and Diner, B. A. (1992) Biochim.
Biophys. Acta, 31, 942-948.
28.Boerner, R. J., Nguyen, A. P., Barry, B. A., and
Debus, R. J. (1992) Biochemistry, 31, 6660-6672.
29.Semin, B. K., and Seibert, M. (2004)
Biochemistry, 43, 6772-6782.
30.Mamedov, F., Sayre, R. T., and Styring, S. (1998)
Biochemistry, 37, 14245-14256.
31.Kolling, D. R. J., Cox, N., Ananyev, G. M., Pace,
R. J., and Dismukes, G. C. (2012) Biophys. J., 103,
313-322.
32.Petrova, I. O., Kurashov, V. N., Semenov, A. Yu.,
and Mamedov, M. D. (2011) J. Photochem. Photobiol. B.,
104, 372-376.
33.Kurashov, V. N., Lovyagina, E. R., Shkolnikov, D.
Y., Solntsev, M. K., Mamedov, M. D., and Semin, B. K. (2009)
Biochim. Biophys. Acta, 1787, 1492-1498.
34.De Las Rivas, J., and Barber, J. (2004)
Photosyn. Res., 81, 329-343.