Phycoerythrin Association with Photosystem II in the Cryptophyte Alga Rhodomonas salina
I. N. Stadnichuk1,a*, T. M. Novikova2, G. S. Miniuk2, V. A. Boichenko3#, Yu. V. Bolychevtseva4, E. S. Gusev1, and E. P. Lukashev5
1Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, 127726 Moscow, Russia2Bach Institute of Biochemistry, Research Center of Biotechnology, Russian Academy of Sciences, 119071 Moscow, Russia
3Institute of Fundamental Problems of Biology of the Russian Academy of Sciences, 142290 Pushchino, Moscow Region, Russia
4Kovalevski Institute of Biology of the Southern Seas, Russian Academy of Sciences, 299011 Sevastopol, Russia
5Lomonosov Moscow State University, Faculty of Biology, 119991 Moscow, Russia
* To whom correspondence should be addressed.
# Deceased.
Received April 9, 2020; Revised May 2, 2020; Accepted May 3, 2020
Cryptophyte algae belong to a special group of oxygenic photosynthetic organisms containing pigment combination unique for plastids – phycobiliproteins and chlorophyll a/c-containing antenna. Despite the progress in investigation of morphological and ecological features, as well as genome-based systematics of cryptophytes, their photosynthetic apparatus remains poorly understood. The ratio of the photosystems (PS)s I and II is unknown and information on participation of the two antennal complexes in functions of the two photosystems is inconsistent. In the present work we demonstrated for the first time that the cryptophyte alga Rhodomonas salina had the PSI to PSII ratio in thylakoid membranes equal to 1 : 4, whereas this ratio in cyanobacteria and higher plants was known to be 3 : 1 and 1 : 1, respectively. Furthermore, it was established that contrary to the case of cyanobacteria the phycobiliprotein antenna represented by phycoerythrin-545 (PE-545) in R. salina was associated only with the PSII, which indicated specific spatial organization of these protein pigments within the thylakoids that did not facilitate interaction with the PSI.
KEY WORDS: cryptophytes, phycobiliproteins, phycoerythrin, chlorophyll a, chlorophyll c, photosystem I, photosystem IIDOI: 10.1134/S000629792006005X
Abbreviations: Chl a, chlorophyll a; Chl c, chlorophyll c; Chl a/c-protein, chlorophyll a/c-protein; PBS, phycobilisome; PE-545, phycoerythrin-545; PSI(II), photosystem I(II).
INTRODUCTION
Flagellate cryptophyte algae (cryptophytes) can inhabit the deep lower layers of phytoplankton due to phycobiliproteins absorbing light in the “green window” of chlorophyll formed by the surface algae of other groups [1-3]. Cryptophytes belong to the group of Chromalveolates composed of various algae containing four-membraned chloroplasts with chlorophyll c (Chl c). Photosynthesis in oxygenic photosynthetic organisms is associated with photooxidation of water and release of oxygen as a result of interaction between the two photosystems. The pigment–protein complexes of the photosystem I (PSI) and the photosystem II (PSII) are localized in thylakoid membranes of cryptophytes similar to other algae and higher plants. They exist as dimers in the PSII and as monomers in the PSI [4, 5]. Antennal complexes, which transfer the absorbed light energy to the photosystems, are very diverse. Three types of antennae are known: (i) the phycobilisome antenna in cyanobacteria and red algae; (ii) the antenna containing Chl a/b in green algae and higher plants; (iii) the antenna containing Chl a/c in numerous groups of Chromophytes. The chlorophyll c of the chlorophyll a/c-proteins is an analog of the chlorophyll b in the Chl a/b-containing proteins. These pigment–protein light-harvesting complexes (LHC) are classified into several protein families. The cryptophyte chlorophyll a/c-complexes belong to the family of CAC-proteins (chlorophyll a/c-proteins) [6, 7]. Common feature of all homologous LHCs is the presence of up to 14-15 chlorophyll molecules in them. The Chl a : Chl b ratio, as well as the ratio Chl a : Chl c in the LHC-families can vary, but Chl a molecules are always predominant [8]. Several chlorophylls c are known: Chl c1, Chl c2, Chl c3, etc. The majority of cryptophyte species have Chl c2 in the cells [6, 9].
The specific feature of cryptophyte algae is that they use a phycobiliprotein antenna to complement the Chl a/c-protein. Cryptophyte phycobiliproteins have been detected neither in cyanobacteria nor in red algae and are unique pigment proteins. According to their color, they are divided into blue phycocyanins (PC) and red phycoerythrins (PE). Seven (or, possibly, eight) protein–pigments of the two-color groups are recognized according to their long-wavelength (nm) absorption maxima: PE-545, PE-555, PE-566, PC-569, PC-612, PC-630, and PC-645 [10, 11]. The resulting color of the proteins and shape of the spectra are determined by different combinations of the chromophores within each phycobiliprotein. Two of six chemically identified phycobilin chromophores, phycocyanobilin and phycoerythrobilin, coincide with the chromophores established for the red algae and cyanobacteria, whereas the other four chromophores are unique [11]. Every phycobiliprotein is a polypeptide heterodimer (α1βα2β), i.e., it contains two different α-subunits and two identical β-polypeptides with the total mass of ~60 kDa. There is one chromophore in the α-polypeptides, and three chromophores in the β-polypeptides; as a result, the dimer contains eight chromophore groups. β-Subunits are encoded in the genome of plastids, but α1 and α2 are nucleus-encoded and are transported across five membranes with the help of signaling peptides into the chloroplast lumen, where they are assembled into dimers. While only two copies of the β-subunit gene [12] exist, the repeated gene duplication has led to development of a large family of α-subunit genes (up to 20 genes); although only two of them are expressed preferentially at every moment depending on the environment [13]. In cyanobacteria and red algae phycobiliptoteins are assembled into phycobilisomes – megacomplexes with the mass of several million Dalton, which include several hundred phycobilin chromophores. In cryptophytes the (α1βα2β)-heterodimers are not assembled into phycobilisomes and are not localized on the stromal surface of the thylakoid membrane similarly to phycobilisomes, but rather in the internal lumenal region of thylakoids [14, 15].
The supramolecular organization of phycobiliproteins and their localization inside the thylakoid lumen are still unclear. The dimers occupy the entire space of the lumen and are likely assembled into cylindrical structures oriented perpendicularly to the thylakoid membrane [14-16]. The currently available data on the structure produced using various approaches do not allow resolving the supramolecular assembly [17]. So far, all attempts to visualize in detail self-assembly of the relatively small dimers (~60 kDa), similar to the self-assembly of phycobilisomes, or to isolate possible protein macrostructures using biochemical approaches failed. Further studies are required to find an answer to this question.
The issue of specific features of Chl-a/c-proteins really comes down to examination of the topography of these proteins in the thylakoid membrane plane. It was found using the single-particle electron microscopy technique that from four to eight Chl-a/c-proteins were attached to the lateral surfaces of the PSII dimers and the PSI monomers [5], markedly repeating the structure of Chl a/b-protein supercomplexes within the PSII and PSI in higher plants and green algae [7]. These data together with various spectral investigations allowed concluding that there was direct transfer of the absorbed energy from the Chl a/c-containing antenna to both photosystems. This makes the question on the role of phycobiliproteins as an antenna system in cryptophytes even more urgent. Migration of energy from phycobiliproteins to Chl a in the cell was revealed using steady state spectroscopy already in the first studies [18], but the possibility of energy migration exclusively to the PSII or also to the PSI is still under examination. The complex of PE-565 and PSII was isolated from the solubilized plastids of the Cryptomonas rufescens alga in the sucrose density gradient [19], and the complex containing both PC-645 and Chl a/c-protein was obtained from the Chroomonas placoidea species [20]. Simultaneous isolation of water-soluble phycobiliproteins and membrane proteins (the PSII or Chl a/c-protein) is challenging and, hence, artifacts cannot be ruled out. The problem is aggravated by the shortage of data on the ratio of two photosystems in plastids of cryptophytes and thus on the fraction of the antennal Chl a within each of them.
There are various ideas regarding the probable association of phycobiliproteins with the photosystems and thus transmission of energy absorbed by phycobiliproteins to the PSI and PSII. The Förster mechanism of energy transfer either direct or mediated through Chl a/c-protein was suggested [21, 22]. Stationary fluorescence and excitation spectra of cells were recorded at room and low temperature, when the fluorescence was associated either only with Chl a of the PSII or with Chl a of both photosystems [23, 24]; superfast fluorescence measurements and global spectral analysis of the cryptophyte algae fluorescence were performed in a wide spectral range [25] that allowed the possibility of redistribution of the absorbed energy between the photosystems.
To elucidate the role of phycobiliproteins in the cryptophyte cells, we compared stationary fluorescence emission parameters, assessed the effect of phycobiliprotein on the degree of photooxidation of the reaction center P700 belonging to the PSI, and recorded the action spectra of both photosystems. In addition, the ratios of PSI, PSII, and antennal complexes were calculated. Algae of the Rhodomonas genus are considered as a suitable model for investigation of cryptophytes and their photosynthetic apparatus. The R. salina species was used in the study, which contains PE-545 together with Chl a/c-protein like many other species of this genus.
MATERIALS AND METHODS
Alga strains and cultivation conditions. The sea microalga Rhodomonas salina (Wislouch) (Cryptomonades) from the collection of the Institute of the Southern Seas Biology, Russian Academy of Sciences, was grown in 100-ml flasks with standard Walney medium at 22°C under continuous illumination with white light (30 µM photons m−2·s−1) with mixing on a magnetic stirrer. Cells were collected by centrifugation at 180g at the end of logarithmic phase of growth on the 5th or 6th day of the growth and used immediately in experiments.
Absorption and fluorescence emission spectra. Absorption of R. salina cells and of PE-545 isolated from them was recorded with a Varian 2300 UV Vis spectrophotometer (USA). To minimize light scattering 2-mm light path cuvettes were used. Fluorescence spectra of the cells were recorded with a Fluorolog-3 spectrofluorimeter (Horiba Jobin Ivon, Japan). Cell samples were placed into 5-mm cuvettes, when experiments were conducted at room temperature, and into glass capillaries with the internal diameter of 2 mm so their optical density at 678 nm was not higher than 0.1, when experiments were conducted at 77 K. Excitation and emission slit width was 3 nm. Spectrum of every sample was recorded 3-5 times, averaged, and after this contribution of the scattered light was subtracted using software supplied with the fluorimeter.
Preparation of PE-545 and thylakoid membranes. Cells washed with 0.01 M K-phosphate buffer (pH 6.5) were subjected to freezing–thawing cycle at –20°C twice causing disintegration of the weak cell wall of cryptophytes and release of PE-545 (as well as other water-soluble proteins) into the buffer solution. The water-insoluble membrane fraction was separated using a table centrifuge followed by salting out the protein–pigment from the supernatant with 60% ammonium sulfate. Next the protein precipitate was dissolved in the same buffer and dialyzed against it. The obtained sample was loaded onto a chromatographic column with an ion-exchange resin DEAE-52 (2 × 11 cm, Whatman, USA) for PE-545 purification. The green precipitate (water-insoluble membrane fraction) was resuspended in 0.1 M Tris-glycine buffer (pH 8.5), and after additional washing and removal of intact cells was used as a preparation of thylakoid membranes.
Reversible photooxidation of P700 was evaluated from the difference of the cell sample absorption at 810 and 870 nm using a dual wavelenght emitter-detector ED-P700 DW (PAM-101, Germany) [26]. After the dark adaptation for 10 min, the sample was illuminated with a halogen lamp KL 1500 (Schott, Germany). Light intensity after interference filters BPF 680/35 or BPF 580/35 (Fotooptik, Russia) and a heat-absorbing filter (Balzers, Lichtenstein) was adjusted to 500 µM photons m–2·s–1; measurements were conducted using 0.3-cm pathlength cuvettes with Chl a samples at concentration 10 µg ml–1.
Photosynthetic action spectra were recorded by both the photo-induced partial release of O2 (the PSII) and by the light-dependent absorption of oxygen (the PSI) on a special device [27] that combined polarography and illumination of cells by monochromatic light flashes in the region from 400 to 720 nm with equal intensity of 0.2 µM cm–2·s–1. Dark periods for recovery of the initial photoactivity of the samples were 40 s. Cell suspension (20 µl) in 50 mM Na-phosphate buffer solution/50 mM KCl (pH 6.8) and optic density of 0.15 (Chl a) was placed onto the surface of a highly sensitive illuminated platinum electrode with an Ag/AgCl reference electrode. Oxygen respiration of the cells was inhibited by illuminating the sample with a weak continuous light at ≥700 nm in order to record the PSII photoactivity. To record the PSI activity, 10 µM DCMU was added to the sample to inhibit the PSII activity, as it was described in detail previously [27, 28]. The spectra of three-five independent cell samples were averaged.
Stoichiometry of pigment–protein complexes and the PSI/PSII ratio were calculated using the known extinction coefficients of PE-545, Chl a and Chl c [29-31]. The PSI and PSII ratio (the ratio of the number of PSI/PSII reaction centers) was determined by summing up the action spectra of the two photosystems and fitting the resulting spectrum to provide minimal deviations from the absorption spectrum of the R. salina cells using iterations with the least squares method in the identified spectral region [28, 32].
RESULTS
Absorption spectra. The absorption spectrum of R. salina cells was typical for the Rhodomonas algae (Fig. 1). The Soret band of Chl a was in the region of 400-440 nm, and of Chl c – at 455 nm; the intensive absorption band of carotenoids including the main one – alloxanthine [3] was at 495 nm. The Chl c absorption maximum in the red region was at 635 nm, the major red band of Chl a absorption was at 676 nm. The absorption spectrum of PE-545 isolated from the R. salina cells (Fig. 1) had a characteristic pattern [33]. The maximum was observed at 545 nm with the closely located shoulder at 565 nm; these bands characterized two types of phycobilins within PE-545, the more short-wavelength phycoerythrobilin and the more long-wavelength dihydrobiliverdin [33].
Fig. 1. Absorption spectra of R. salina cells (1); of PE-545 isolated from the cells (2), of the thylakoid membranes (3), and the combined spectrum (4) (the spectrum 2 + the spectrum 3) modeling the spectrum (1) of the cells.
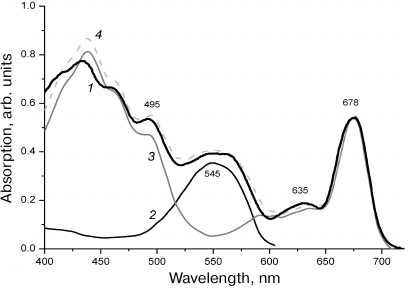
The region of the long-wavelength shoulder in the spectrum of the intact cells assigned to the absorption of PE-545 is markedly broaden due to overlap with the satellite bands of Chl c and Chl a, which are also pronounced in the spectrum of the Chl a/c-protein [8] isolated from the thylakoids and in the action spectrum of the PSI (see further). The bands observed in the spectrum of thylakoid membranes of R. salina repeat the shape of the absorption spectrum of intact cells, except the region of PE-545 absorption (Fig. 1).
Fluorescence spectra of R. salina cells (Fig. 2a) at room temperature were recorded with excitation at two wavelengths: 440 nm (Soret bands of Chl a and Chl c) and 525 nm (PE-545). In the first case, the spectrum had one intensive band of the PSII at 685 nm. More short-wavelength fluorescence, which could theoretically belong to Chl c was not observed, similar to the absence of Chl b fluorescence in higher plants. Its absence implies virtually 100% efficiency of energy transfer from Chl c to Chl a.
Fluorescence excitation at 525 nm, in the region of PE-545 absorption, also resulted in intensive emission of Chl a with the maximum at 685 nm coinciding fully with the previous spectrum, which implied energy transfer from the phycobiliprotein antenna (Fig. 2a). The observed short-wavelength band of the lower intensity at 588 nm is known to belong to PE-545 itself [10, 34].
Fig. 2. Fluorescence emission spectra of R. salina cells recorded at room temperature (a) and at 77 K (b). Fluorescence excitation at 440 nm (1) and at 525 nm (2). The spectra are normalized to emission maxima.
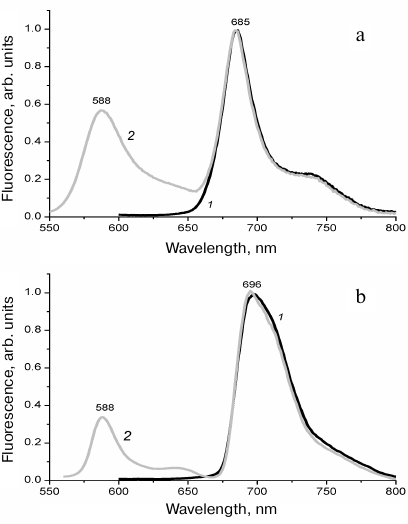
No long-wavelength fluorescence of the PSI was observed in the spectra of R. salina recorded at low temperature (77 K). The difference with the spectra recorded at room temperature is manifested by a minor red shift of the Chl a maximum from 685 to 696 nm (Fig. 2b). The changes were too small to identify reliably two photosystems in the spectra. Individual bands of the PSI and PSII cannot be discriminated, in contrast to the low-temperature fluorescence of higher plants, for example, therefore, the probability of PE-545 participation in the PSI activity does not become clearer when stationary fluorescence spectra are used.
Photoinduced oxidation of P700. The primary charge photoseparation in the P700 reaction center indicates the energy transfer of the absorbed light energy to the PSI. To reveal this transfer, it is necessary to compare the degree of photooxidation of P700 at two wavelengths: the 680 nm light directly excites Chl a, whereas at 580 nm the antennal complex of PE-545 is excited preferentially. Comparison of the degree of reversible photooxidation of P700 indicated that in the latter case the signal decreased twofold in comparison with the direct excitation of Chl a within the PSI complex (Fig. 3). According to the absorption spectrum (Fig. 1), 75% of the 580-nm light was absorbed by PE-545 in the cells.
Fig. 3. Photoinduced changes in the absorption of the reaction center P700 in the R. salina cells illuminated for 30 s by 680-nm light (1) or 580-nm light (2) in the absence of diuron. The arrows ↑ and ↓ show the light switching on and off.
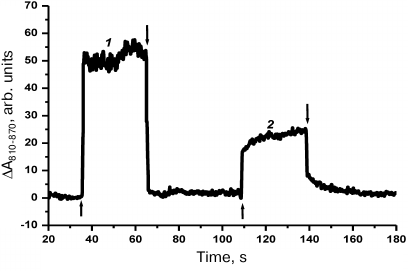
One can assume that the twofold decrease in the signal (Fig. 3) indicates a small contribution of PE-545 to functioning of the PSI. The drop in intensity was accompanied by slowing down of the P700 photooxidation kinetics (regions between the arrows ↑ and ↓ in Fig. 3). This indicated an increase in the light absorption by the PSII at 580 nm that had to be accompanied by the increase of the linear electron flow from the PSII to the PSI. “Feeding” of the PSI by additional negative charges was manifested by the decrease in the P700 photooxidation. As a result, in the presence of indication of the PE-545 association with the PSII it was impossible to tell with certainty whether the decrease in the P700 photooxidation was caused by insufficient energy migration from PE-545 to the PSI, by specific features of the two photosystems interaction, or by any other reason.
Photosynthetic action spectra. The action spectra were recorded under conditions of absence of spectral data of the PSI activity during absorption of light by the PSII and vice versa. Therefore, the pattern of the action spectrum of each photosystem repeats the spectrum of its absorption not influenced by the presence of the other photosystem in the thylakoids [27].
Fig. 4. a) Photosynthetic action spectra of the PSI (1) and PSII (2) registered for R. salina cells. The spectra are normalized to the red absorption peak of Chl a at 676-678 nm. Inset: 3) difference between the spectra (2) and (1); 4) absorption spectrum of PE-545; b) absorption spectrum of the R. salina cells (1), and the spectrum modeled by the sum of the action spectra of the PSI and PSII presented in Fig. 4a and taken at the ratio of 40 and 60% of the chlorophyll absorption maximum (2).
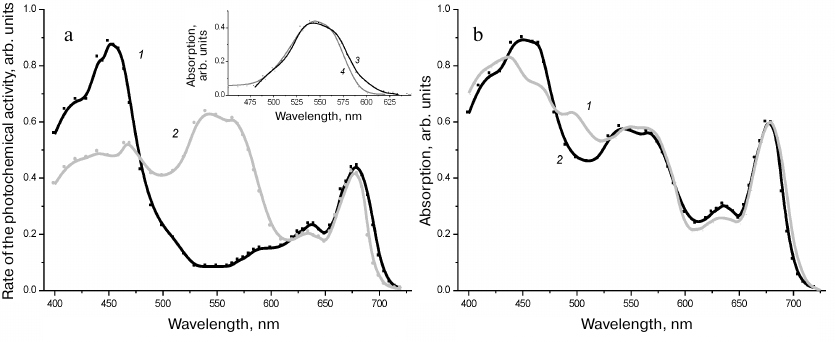
The photosynthetic action spectrum of the PSII in the short wavelength region from 400 to 470 nm comprised superposition of the bands belonging to Chl a, Chl c, and carotenoids. In the orange-red region a very intensive band at 545 nm was observed together with the maximum of Chl a at 676 nm and the less intensive maximum of Chl c at 636 nm, which could be clearly assigned to PE-545 with the long-wavelength shoulder at 555-575 nm contributed to the Chl a/c-protein (Fig. 4a). The photosynthetic action spectrum of the PSI (Fig. 4a) was characterized by all the bands belonging to Chl a and Chl c and identified in both the absorption spectrum of the cells and the action spectrum of the PSII, but with two specific features differentiating it from the PSI spectrum. The greater relative intensity of the blue band of Chl c at 460 nm in comparison with the PSII spectrum means also the greater fraction of the Chl a/c-protein in the composition of PSI [31]. However, the main specific feature of the PSI action spectrum was the complete absence of spectral features of PE-545 that demonstrated very clearly association of the PE-4545 only with the PSII. This conclusion was strongly corroborated by the difference spectrum of these two curves (Fig. 4a, inset) pattern of which resembled closely the absorption spectrum of PE-545 presented in Fig. 1.
PSI/PSII ratio. The ratio of the two photosystems in chloroplasts is not a trivial task; electrophoresis of thylakoid membranes, the atrasine approach for detecting the PSII activity, and some auxiliary techniques including photooxidation of P700, have been used with varying reliability to address it. We used the most accurate of the methods – modeling of the absorption spectrum by the sum of action spectra, because the sum of two spectra taken in an adequate proportion must repeat its pattern and thus establish the ratio of the two photosystems. As in the case of phycobilisomes in cyanobacteria [28], the significant range for calculations of the sum spectra is from 500 to 700 nm, because in the spectral region 400-500 nm some carotenoids of the photosystems cannot transfer 100% of the energy to Chl a making impossible to obtain correct data. As a result the ratio of the two photosystems (PSI/PSII), more accurately, of their reaction centers, was established as 1 : 4 (Fig. 4b). This result was obtained from comparison of the amplitudes of the red bands at 676-678 nm in the action spectra, the intensity of which was proportional to the Chl a content in the core complexes of the PSI and PSII. The ratio of the reaction centers was obtained with consideration that the PSI (96 chlorophylls [35]) has the 2.6 times greater chlorophyll antenna than the PSII (36 chlorophyll molecules [36]). As mentioned above, dimers of the PSII and monomers of the PSI are real structures of the thylakoid membrane. Thus, two dimers of the PSII per one monomer of the PSI are present in the R. salina thylakoids.
Stoichiometry of antennal pigment–protein complexes of the PSI and PSII. To elucidate specific features of the molecular organization of the pigment apparatus in cryptophytes it was important to find out the size of the phycoerythrin antenna, i.e., the number of PE-545 dimers per one reaction center of the PSII. Modeling of the absorption spectrum of the R. salina cells by the sum of the thylakoid and PE absorption spectra showed that it was possible to assess contribution of the PE absorption to the spectrum and to determine its ratio with Chl a (Fig. 1). However, calculations were complicated by the fact that part of Chl a in the thylakoids was in the composition of the Chl a/c-protein. Therefore, the data on stoichiometry of the antennal complex of the Chl a/c-protein and the core complexes of the PSII and PSI are required in addition to the PE-545 data. They can be obtained by evaluating contributions of every component to the absorption maxima of other pigments with the use of molar extinction coefficients maxima (PE-545 at 545 nm; Chl c at 636 nm, and Chl a at 676-678 nm). General method of the calculations has been described in detail in [28].
If one takes absorption of the PE545 in relative values to be 1, it can be deduced from the Fig. 1 that for PE-545:
PE545 = 1.00; PE636 = 0.00, and PE676-8 = 0.00 (1).
Relative absorptions of Chl a obtained from the data on the pigment in solution and in the core complex of the PSII are also known [28]:
Chl a676-8 = 1.00; Chl a636 = 0.26, and Chl a545 = 0.1 (2);
similarly, absorption rates for Chl c are [31]:
Chl c636 = 1.00; Chl c676-8 = 0.00, and Chl c545 = 0.14 (3).
The question is how to obtain molar ratio of the pigments based on the ratios of intensities of the absorption bands (1)-(3), and how to test correctness of these calculations. Molar extinction coefficient of PE-545 {PE545} is 730 mM–1·cm–1 assuming that molecular mass of the (α1βα2β)-dimer is 58 kDa [29]. Molar extinction coefficient of Chl a in vivo is {Chl a676} = 64 mM–1 ·cm–1 [30], which is lower than the extinction coefficient of Chl a in different solvents [31]. For Chl c the extinction coefficient in the red band is twice as low as the Chl a coefficient [31] that brings the value for the pigment to {Chl c636} = 32 mM–1·s–1. Obviously, the PE-545 contribution to the absorption at 636 nm, where the maximum of the Chl a/c-protein band is located, and especially at 676-678 nm (the red maximum of Chl a) according to (1) can be disregarded; according to (3) analogously, the Chl c contribution to the Chl a in its red maximum is neglected too that minimizes the stoichiometric calculations.
Determination of the molar ratios of the Chl a/c-protein complex and core complexes of the photosystems is the easiest. Taking into consideration the above-mentioned twofold lower extinction coefficient of Chl c in comparison with Chl a and the ratio of the amplitudes (2) and (3), the molar fraction {Chl c} relative to the molar fraction {Chl a} for the absorption spectrum D measured in optical density units or for the corresponding photosynthetic action spectra is determined by the formula:
{Chl c}/{Chl a} = 2(D636 – 0.26D676-8)/64 = 0.16(D636 – 0.26D676-8) (4).
According to (4), the {Chl a}/{Chl c} ratio for thylakoids of the investigated R. salina species (Fig. 1) was 4.1. Extraction of the cellular Chl a and Chl c with 80% acetone [8] resulted in the same ratio of pigments in the extract (data not presented). The calculation result is in full agreement with the ratio 4 : 1 that was found for the closely related species R. lens after extraction and pigment separation by chromatography [8]. The obtained data provides a strong support to the suggested approach for calculating the Chl a/Chl c ratio in the cells of R. salina, R. lens and other species of the PE-545 containing cryptophyte algae.
Using the obtained data and recorded action spectra, one may attempt calculating stoichiometry of pigment–protein complexes for both photosystems. Composition of the Chl a/c-protein [8] similarly to the composition of Chl a/b-protein [37], corresponds to the simultaneous presence of 14 molecules of two pigments, six molecules of Chl c, and eight molecules of Chl a bound to each apoprotein molecule. As in the absorption spectra, the red maximum amplitude of the Chl a action spectrum at 676-678 nm is due to contributions of two components: absorption of own Chl a in the core complex of the PSI or PSII and absorption of Chl a molecules within the Chl a/c-proteins attached to every core complex in the thylakoid membrane [5]. Taking into account the above-mentioned ratio (6 : 8) of the pigments in the Chl a/c-protein and after subtracting from the red maximum amplitude of Chl a the absorption part of the chlorophyll molecules in Chl a/c-protein, we obtained the ratio of 25 Chl c molecules per every 72 molecules of Chl a comprising the PSII dimer. Similar calculations performed for the photosynthetic action spectrum of the PSI demonstrated the presence of 35-36 Chl c molecules per every 96 molecules of Chl a (the PSI monomer). These ratios, rounded to the nearest tenths, allowed us to conclude that every dimer of PSII was most likely associated with four pigment–protein complexes of Chl a/c-protein(s) and that the PSI monomers in the thylakoid membrane were in contact with six Chl a/c-antennal complexes, on average. These calculations results are in full agreement with the electron microscopy data [5].
The obtained photosynthetic action spectrum of the PSII (Fig. 4a) allowed us to perform similar calculations of the molar ratio between chlorophyll dimers of the PSII and PE-545. Since every dimer of the PSII (72 molecules of Chl a) is bound, on average, with four Chl a/c-proteins (32 molecules of Chl a), the measured amplitude of the Chl a band at 676 nm in the action spectrum for the PSII has to be decreased by 40% in the calculations. Concurrently, because of the contribution of the Chl c and Chl a absorption to the 545 nm band, its intensity, according to (2) and (3), has to be decreased by ~20%. Taking into account these spectral corrections and molar extinction coefficients, every dimer of the PSII in the R. salina cells obtains energy from nine dimers of PE-545, on average. In comparison with cyanobacteria, this value seems not too high. Indeed, a typical phycobilisome contacting with the PSII dimer in cyanobacteria contains 250-300 phycobilin chromophores [28]. The presence of up to nine dimers in our case indicates the binding of only of 8 × 9 = 72 phycobilin chromophores with the PSII dimer, which is fourfold less than the “phycobilisomal” variant. This difference in the phycobiliprotein antenna size is partly compensated in cryptophytes by the presence of another antenna, Chl a/c-protein [38].
DISCUSSION
The fact of migration of the absorbed light energy from phycobiliproteins to the PSII in cryptophytes, as well as from phycobilisomes in cyanobacteria and red algae was never doubted. It was established in numerous studies over many years that phycobilisomes transfer energy to both the PSII and PSI [28]. A possible analogy with phycobilisomes led to suggestion that such option should also exist in the cryptophyte algae [39]. Stationary fluorescence emission and fluorescence excitation spectra indicated preferential [23] or exclusive [24] interaction of phycobiliproteins with the PSII. Time-resolved fluorescence technique and global spectral analysis based on multiple measured kinetics and theoretical models of the excitation energy transfer considered the migration of energy to both photosystems to be very likely [25]. An extreme sensitivity of cryptophyte algae as flagellate microorganisms to the changes of cultivation conditions and of sample preparation procedures makes artifacts very probable. In the works [25, 34] this was manifested by high intensity of intrinsic fluorescence of phycobiliproteins that was markedly higher that the emission of Chl a as the cell component, whereas typically ([23, 24] and the present work, Fig. 2) the chlorophyll fluorescence is predominant. Among kinetic components of excitation, data appear with the lifetimes specific for phycobiliproteins in solution [25, 34], but not in the cell, where energy is transferred from the antenna to the photosystems. These data indicate weakening of the contacts of phycobiliproteins with the thylakoid membrane and loss of the chloroplast intactness, which leads in the case of [25, 34] to appearance of additional spectral components and possible incorrect conclusions. Preparative isolation of phycobiliproteins together with membrane pigment complexes with the help of biochemical techniques is challenging due to their different hydrophobicity [19, 20]. Under these conditions, it was important to use a non-invasive spectral approach to examine association of phycobiliprotein with photosystems, and it was realized in our work. Three approaches were employed: (i) low-temperature cell fluorescence emission spectra; (ii) photooxidation of the reaction center P700; and (iii) photosynthetic action spectra of the photosystems. The first approach provided no answer as no long-wavelengths fluorescence was observed in R. salina. The second approach only showed restrictions in the energy migration from PE-545 to the PSI because of overlap of the absorption spectra of PE-545 and Chl a/c-protein. Only the third approach – the recording of action spectra of photosynthetic activities, which has not been used earlier for investigation of the pigment apparatus in cryptophytes, produced a clear result indicating the absence of PE-545 association with the PSI.
The action spectrum for the PSII of the R. salina contains a high intensity maximum of PE-545 similar to the bands of phycobilisomes in the spectra of the PSII in cyanobacteria and red algae [28]. However, in the photosynthetic action spectrum of the PSI the band of PE-545 was not observed (Fig. 4a), therefore, affinity of the phycobiliprotein antenna for the PSII is selective. If the contact between the pigment–protein complexes of the two photosystems in the thylakoid membrane of R. salina existed, part of the energy from PE-545 would reach the PSI owing to the PSII mediation. Such contact that leads to the energy “spillover” between the photosystems is not ruled out for the chloroplasts of higher plants [40]. Since no such events were observed in cryptophytes, the PSI and PSII interact on the level of electron transport chain, rather than on the level of energy migration. Thus, we can conclude that the phycobiliproteins of cryptophyte algae, which unlike phycobilisomes are in a finely dispersed state of (α1βα2β)-dimers, contact only with the PSII.
The diffusion coefficients of phycobiliproteins in the lumen of chloroplasts in cryptophytes indicate that their mobility is decreased in comparison with the mobility, which could be expected in the absence of restrictions [41]. The size of dimers, according to X-ray crystallography data, is 4 × 6 × 7 nm [41], that is nearly two orders of magnitude lower than the average size of phycobilisomes in cyanobacteria, but the diffusion coefficients become comparable after the size correction [41]. By analogy with phycobilisomes anchored on the outer side of the thylakoid membrane of cyanobacteria, this comparison can suggest binding of PE-545 to certain sites on the lumenal surface of the membranes. The existance of restrictions of the dimer diffusion is in good agreement with our data on the selective binding of PE-545 to the PSII.
A significant part of the lumenal membrane surface of the PSII complexes is occupied by a chlorophyll-free water-oxidizing complex [36] that in cryptophytes limits the area of the pigment contact with the phycobiliprotein dimers. Therefore, a priori there is a probability of a direct contact or contact mediated by Chl a/c-proteins that interact laterally with the PSII dimer thus increasing the total surface of interaction. However, the non-photochemical quenching realized in cryptophytes through Chl a/c-protein without involvement of phycobiliproteins indicates spatial disconnection of the two antennal complexes in the thylakoids [42]. The findings leave the problem open for the further studies. No less interesting is the opposite question on mechanisms or structures that prevent joining of PE-545 in our case to the PSI surface. Since the Chl a/c-proteins are also components of the PSI, this seems adding to its similarity to the PSII, but does not help to resolve the problem. It can be suggested that the positive surface charge of the PSI required for the interaction with plastoquinone pool as a participant of the cyclic electron transport might play a role in this.
Selectivity of phycobiliproteins towards the PSII indicates their self-assembly that creates a supramolecular architecture in the internal space of the thylakoid. The self-assembly of phycobilisomes is facilitated by the so-called linker, or binding, proteins, but no linkers were found for the (α1βα2β)-dimers either during their isolation or genome analysis. They may be difficult to recognize if they are not homologous to the linkers of phycobilisomes; and that leaves the question open. The shape of (α1βα2β)-dimers is somewhat like a saucer. There is a probability of virtual packing of such “saucers” in cylindric structures mentioned in the literature on electron microscopy [14-16]. The lumen cross section dimensions allow accommodating cylinder-shaped blocks formed by four, or maximum, five “saucers”. In this case two cylinders of PE-545 dimers must interact with each PSII dimeric complex inside the lumen, which does not contradict our calculated data.
The questions on energy transfer from the antennal complexes to the PSI and PSII complexes and the fraction of each photosystem in the thylakoids are closely related. In higher plants, the ratio of the two photosystems is close to 1 : 1. In cyanobacteria, the content of the PSI is two-three-fold higher than the content of the PSII [28]. Therefore, the ratio of PSI/PSII equal to 1 : 4 established by us for R. salina suggests specific features of electron transfer in the thylakoids of cryptophyte algae. Due to the linear transport between the photosystems, the light stage of photosynthesis results in production of ATP and NADPH. The cyclic electron transfer in the PSI is sufficient only for synthesis of ATP [43]. Therefore, the increased fraction of the PSII in cryptophytes indicates the higher biosynthesis of NADPH in comparison with higher plants and cyanobacteria. It should be noted that this issue is poorly understood for all other photosynthesizing flagellates, although such type of movement undoubtedly must have its impact on the processes of energy supply. Further studies will help to elucidate what biochemical features of the cryptophyte cells are associated with these differences.
Funding. The work was financially supported in part of T. M. Novikova and G. S. Miniuk by the State Budget Project No. 0828-2020-0004 (AAAA-A18-118021350003-6), Institute of the Southern Seas.
Conflict of interest. The authors declare no conflict of interest.
Compliance with ethical norms. This article does not contain any studies with human participants or animals performed by any of the authors.
REFERENCES
1.Krasnova, E. D., Pantyulin, A. N., Matorin, D. N.,
Todorenko, D. A., Belevich, T. A., Milyutina, I. A., and Voronov, D. A.
(2014) Blooming of the cryptophyte alga Rhodomonas sp.
(Cryptophyta, Pyrenomodaceae) in the redox-zone of reservoirs separated
from the White Sea, Mikrobiologiya, 83, 346-354,
doi: 10.7868/S0026365614030100.
2.Mitrofanova, E. Y. (2015) Chroomonas acuta
Uterm. (Cryptophyta) in the Lake Teletskoe (Altai, Russia),
Turczaninowia, 18, 96-104,
doi: 10.14258/turczaninowia.18.2.10.
3.Hoef-Emden, K., and Archibald, J. M. (2017)
Cryptophyta (Cryptomonads), in Handbook of the Protists
(Archibald, J. M. et al., eds.) Springer International Publishing AG
2017, 851-891, doi: 10.1007/978-3-319-28149-0_35.
4.Janssen, J., and Rhiel, E. (2008) Evidence of
monomeric photosystem I complexes and phosphorylation of chlorophyll
a/c-binding polypeptides in Chroomonas sp. strain
LT (Cryptophyceae), Intern. Microbiol., 11, 171-178,
doi: 10.2436/20.1501.01.57.
5.Kereïche, S., Kouřil, R., Oostergetel,
G. T., Fusetti, F., Boekema, E. J., Doust, A. B., Van der Weij-de Wit,
C. D., and Dekker, J. P. (2008) Association of chlorophyll
a/c2 complexes to photosystem I and photosystem II in
the cryptophyte Rhodomonas CS24, Biochim. Biophys. Acta,
1777, 1122-1128, doi: 10.1016/j.bbabio.2008.04.045.
6.Hoffman, G. E., Sanchez-Puerta, M. V. S., and
Delwiche, C. F. (2011) Evolution of light-harvesting complex proteins
from Chl c-containing algae, BMC Evol. Biol., 11,
101, doi: 10.1186/1471-2148-11-101.
7.Neilson, J. A. D., and Durnford, D. G. (2010)
Structural and functional diversification of the light-harvesting
complexes in photosynthetic eukaryotes, Photosynth. Res.,
106, 57-71, doi: 10.1007/s11120-010-9576-2.
8.Ingram, K., and Hiller, R. G. (1983) Isolation and
characterization of a major chlorophyll a/c2
light-harvesting protein from a Chroomonas species
(Cryptophyceae), Biochim. Biophys. Acta, 722,
310-319, doi: 10.1016/0005-2728(83)90078-6.
9.Schimek, C., Stadnichuk, I. N., Knaust, R., and
Wehrmeyer, W. (1994) Detection of chlorophyll c1 and
magnesium-2,4-divinylpheoporphyrin A5 monomethylester in
cryptophytes, J. Phycol., 30, 621-627,
doi: 10.1111/j.0022-3646.1994.00621.x.
10.Hill, D. R., and Rowan, K. S. (1989) The
biliproteins of the cryptophyceae, Phycologia, 28,
455-463, doi: 10.2216/i0031-8884-28-4-455.1.
11.Glazer, A. N., and Wedemayer, G. J. (1995)
Cryptomonad biliproteins: an evolutionary perspective, Photosynth.
Res., 46, 93-105, doi: 10.1007/BF00020420.
12.Broughton, M. J., Howe, C. J., and Hiller, R. G.
(2006) Distinctive organization of genes for light-harvesting proteins
in the cryptophyte alga Rhodomonas, Gene, 369,
72-79, doi: 10.1016/j.gene.2005.10.026.
13.Kieselbach, T., Cheregi, O., Green, B. R., and
Funk, C. (2018) Proteomic analysis of the phycobiliprotein antenna of
the cryptophyte alga Guillardia theta cultured under different
light intensities, Photosynth. Res., 135, 149-163,
doi: 10.1007/s11120-017-0400-0.
14.Ludwig, M., and Gibbs, S. P. (1989) Localization
of phycoerythrin at the lumenal surface of the thylakoid membrane in
Rhodomonas lens, J. Cell Biol., 108, 875-884,
doi: 10.1083/jcb.108.3.875.
15.Spear-Bernstein, L., and Miller, K. R. (1989)
Unique location of the phycobiliprotein light-harvesting pigment in the
cryptophyceae, J. Phycol., 25, 412-419,
doi: 10.1111/j.1529-8817.1989.tb00245.x.
16.Mörschel, E., and Wehrmeyer, W. (1979)
Elektronenmikroskopische feinstrukturanalyse von nativen
biliproteidaggregaten und deren räumliche ordnung, Ber. Dtsch.
Bot. Ges., 92, 393-402,
doi: 10.1111/j.1438-8677.1979.tb03286.x.
17.Vesk, M., Dwarte, D., Fowler, S., and Hiller, R.
G. (1992) Freeze fracture immunocytochemistry of light-harvesting
pigment complexes in a cryptophyte, Protoplasma, 170,
166-176, doi: 10.1007/BF01378791.
18.Haxo, F. T., and Fork, D. C. (1959)
Photosynthetically active accessory pigments of cryptomonads,
Nature, 184, 1051-1052, doi: 10.1038/1841051a0.
19.Lichtlé, C., Duval, J. D., and Lemoine, Y.
(1987) Comparative biochemical, functional and ultrasructural studies
of photosystem particles from a Cryptophyceae Cryptomonas
rufescens: isolation of an active phycoerythrin particle,
Biochim. Biophys. Acta, 894, 76-90,
doi: 10.1016/0005-2728(87)90214-3.
20.Chen, M., Li, S. H., and Sun, L. (2007) A novel
phycocyanin–Chl a/c2–protein complex
isolated from chloroplasts of Chroomonas placoidea, Chinese
Chem. Lett., 18, 1374-1378,
doi: 10.1016/j.cclet.2007.09.025.
21.MacColl, R., and Berns, D. S. (1978) Energy
transfer studies on cryptomonad biliproteins, Photochem.
Photobiol., 27, 343-349,
doi: 10.1111/j.1751-1097.1978.tb07610.x.
22.Mimuro, M., Tamai, N., Murakami, A., Watanabe,
M., Erata, M., Watanabe, M. M., Tokutomi, M., and Yamazaki, T. (1998)
Multiple pathways of excitation energy flow in the photosynthetic
pigment system of a cryptophyte, Cryptomonas sp. (CR-1),
Phycol. Res., 46, 155-164,
doi: 10.1111/j.1440-1835.1998.tb00108.x.
23.Bruce, D., Biggins, J., Steiner, T., and Thewalt,
M. (1986) Excitation energy transfer in the cryptophytes. Fluorescence
excitation spectra and picosecond time-resolved emission spectra of
intact algae at 77 K, Photochem. Photobiol., 44, 519-525,
doi: 10.1111/j.1751-1097.1986.tb04702.x.
24.Lichtlé, C., Jupin, C. H., and Duval, I.
C. (1980) Energy transfer from PSII to PSI in Cryptomonas
rufescens (Cryptophyceae), Biochim. Biophys. Acta,
591, 104-112, doi: 10.1016/0005-2728(80)90224-8.
25.Van der Weij-de Wit, C. D., Doust, A. B., Van
Stokkum, I. H. M., Dekker, J. P., Wilk, K. E., Curmi, P. M. G.,
Scholes, G. D., and Van Grondelle, R. (2006) How energy funnels from
the phycoerythrin antenna complex to photosystem I and photosystem II
in cryptophyte Rhodomonas CS24 cells, J. Phys. Chem. B,
110, 25066-25073, doi: 10.1021/jp061546w.
26.Schreiber, U., Klughammer, C., and Neubauer, C.
(1988) Measuring P700 absorbance changes around 830 nm with a new
type of pulse modulation system, Z. Naturforsch., 43,
686-698, doi: 10.1515/znc-1988-9-1010.
27.Boichenko, V. A. (1998) Action spectra and
functional antenna sizes of photosystems I and II in relation to the
thylakoid membrane organization and pigment composition, Photosynth.
Res., 58, 163-174, doi: 10.1023/A:1006187425058.
28.Rakhimberdieva, M., Boichenko, V., Karapetyan,
N., and Stadnichuk, I. (2001) Interaction of phycobilisomes with
photosystem II dimers and photosystem I monomers and trimers in the
cyanobacterium Spirulina platensis, Biochemistry,
40, 15780-15788, doi: 10.1021/bi010009t.
29.MacColl, R., Berns, D. S., and Gibbons, O. (1976)
Characterization of cryptomonad phycoerythrin and phycocyanin, Arch.
Biochem. Biophys., 177, 265-275,
doi: 10.1016/0003-9861(76)90436-7.
30.Rögner, M., Mühlenhoff, U., Boekema, E.
J., and Witt, H. (1990) Mono-, di- and trimeric PSI reaction center
complexes isolated from the thermophilic cyanobacterium
Synechococcus sp.: size, shape and activity, Biochim.
Biophys. Acta, 1015, 415-424,
doi: 10.1016/0005-2728(90)90074-E.
31.Jeffrey, S. W., and Humphrey, G. F. (1975) New
spectrophotometric equations for determining chlorophylls a,
b, c1 and c2 in higher
plants, algae and natural phytoplankton, Biochem. Physiol.
Pflanzen, 167, 191-194,
doi: 10.1016/S0015-3796(17)30778-3.
32.Boichenko, V. A., Pinevich, A. V., and
Stadnichuk, I. N. (2007) Association of chlorophyll a/b-binding
Pcb proteins with photosystems I and II in Prochlorothrix
hollandica, Biochim. Biophys. Acta, 1767, 801-806,
doi: 10.1016/j.bbabio.2006.11.001.
33.Doust, A. B., van Stokkum, I. H. M., Larsen, D.
S., Wilk, K. E., Curmi, P. M. G., van Grondelle, R., and Scholes, G. D.
(2005) Mediation of ultrafast light-harvesting by a central dimer in
phycoerythrin 545 studied by transient absorption and global analysis,
J. Phys. Chem. B, 109, 14219-14226,
doi: 10.1021/jp051173j.
34.Van der Weij-De Wit, C. D., Doust, A. B., Van
Stokkum, I. H. M., Dekker, J. P., Wilk, K. E., Curmi, P. M. G., and Van
Grondelle, R. (2008) Phycocyanin sensitizes both photosystem I and
photosystem II in cryptophyte Chroomonas CCMP270 cells,
Biophys. J., 94, 2423-2433,
doi: 10.1529/biophysj.107.113993.
35.Jordan, P., Fromme, P., Witt, H. T., Klukas, O.,
Saenger, W., and Krauß, N. (2001) Three dimensional structure of
cyanobacterial photosystem I at 2.5 Å resolution, Nature,
411, 909-917, doi: 10.1038/35082000.
36.Ferreira, K. N, Iverson, T. M., Maghlaoui, K.,
Barber, J., and Iwata, S. (2004) Architecture of the photosynthetic
oxygen-evolving center, Science, 303, 1831-1838,
doi: 10.1126/science.1093087.
37.Kühlbrandt, W., Wang, D. N., and Fujiyoshi,
Y. (1994) Atomic model of plant light-harvesting complex by electron
crystallography, Nature, 367, 614-621,
doi: 10.1038/367614a0.
38.Cunningham, B. R., Greenwold, M. J., Lachenmyer,
E. M., Heidenreich, K. M., Davis, A. C., Dudycha, J. L., and
Richardson, T. L. (2019) Light capture and pigment diversity in marine
and freshwater cryptophytes, J. Phycol., 55, 552-564,
doi: 10.1111/jpy.12816.
39.Doust, A. B., Wilk, K. E., Curmi, P. M. G., and
Scholes, G. D. (2006) The photophysics of cryptophyte light-harvesting,
J. Photochem. Photobiol. A, 184, 1-17,
doi: 10.1016/j.jphotochem.2006.06.006.
40.Yokono, M., and Akimoto, S. (2018) Energy
transfer and distribution in photosystem super/megacomplexes of plants,
Curr. Opin. Biotechnol., 54, 50-56, doi:
10.1016/j.copbio.2018.01.001.
41.Mirkovic, T., Wilk, K. E., Curmi, P. M. G., and
Scholes, G. D. (2009) Phycobiliprotein diffusion in chloroplasts of
cryptophyte Rhodomonas CS24, Photosynth. Res.,
100, 7-17, doi: 10.1007/s11120-009-9412-8.
42.Kuthanová Trsková, E., Bína,
D., Santabarbara, S., Sobotka, R., Kana, R., and Belgio, E. (2019)
Isolation and characterization of CAC antenna proteins and photosystem
I supercomplex from the cryptophytic alga Rhodomonas salina,
Physiol. Plant., 166, 309-319, doi:
10.1111/ppl.12928.
43.Dann, M., and Leister, D. (2019) Evidence that
cyanobacterial Sll1217 functions analogously to PGRL1 in enhancing
PGR5-dependent cyclic electron flow, Nat. Commun., 10,
5299, doi: 10.1038/s41467-019-13223-0.